
Sigma-2 Receptor/TMEM97 and PGRMC-1 Increase the Rate of Internalization of LDL by LDL Receptor through the Formation of a Ternary Complex

By Aladdin Riad, Chenbo Zeng, Chi-Chang Weng, Harrison Winters, Kuiying Xu, Mehran Makvandi, Tyler Metz, Sean Carlin, and Robert H. Mach
Excerpt from the article published in Scientific Reports 8, 16845, 15 November 2018, DOI: https://doi.org/10.1038/s41598-018-35430-3
Editor’s Highlights
- The disruption of lipoprotein trafficking may also be responsible for the mechanism of action of sigma-2 receptors (TMEM97/σ2R) antagonists in preventing Aβ oligomer synaptic dysfunction and Aβ-induced neurotoxicity.
- Apolipoprotein-E (Apo-E) is the main source of cholesterol in brain, which is required for axonal growth and synaptogenesis.
- Apo-E is synthesized in astrocytes and microglia and released into the extracellular space where it is taken up in neurons by LDLR and a second lipoprotein receptor, LDL receptor related protein 1 (LRP1).
- The uptake of LDL by LDLR is increased by the formation of a complex with both PGRMC1 and TMEM97.
- The involvement of TMEM97/σ2R in the internalization and trafficking of cholesterol via LDL is consistent with our previous characterization of the σ2R is upregulated in proliferating versus quiescent breast cancer cells.
- A disruption of cholesterol biosynthesis has also been proposed as a potential mechanism for treating cancer.
Abstract
CRISPR/Cas gene studies were conducted in HeLa cells where either PGRMC1, TMEM97 or both proteins were removed via gene editing. A series of radioligand binding studies, confocal microscopy studies, and internalization of radiolabeled or fluorescently tagged LDL particles were then conducted in these cells. The results indicate that PGRMC1 knockout (KO) did not reduce the density of binding sites for the sigma-2 receptor (σ2R) radioligands, [125I]RHM-4or [3H]DTG, but a reduction in the receptor affinity of both radioligands was observed. TMEM97 KO resulted in a complete loss of binding of [125I]RHM-4 and a significant reduction in binding of [3H]DTG. TMEM97 KO and PGRMC1 KO resulted in an equal reduction in the rate of uptake of fluorescently-tagged or 3H-labeled LDL, and knocking out both proteins did not result in a further rate of reduction of LDL uptake. Confocal microscopy and Proximity Ligation Assay studies indicated a clear co-localization of LDLR, PGRMC1 and TMEM97. These data indicate that the formation of a ternary complex of LDLR-PGRMC1-TMEM97 is necessary for the rapid internalization of LDL by LDLR.
Introduction
The sigma receptors represent a family of proteins whose function in cell biology is poorly understood. The current classification lists two types of sigma receptors, sigma-1 (σ1) and sigma-2 (σ2) receptors (R)1,2,3. The sigma receptors were identified in 1976 and were initially classified as a member of the opiate receptors4. Subsequent studies revealed that they represented a discrete family of receptors. For many years, the sigma receptors were described pharmacologically through the binding of the radioligands [3H](+)−pentazocine and [3H]DTG. [3H](+)−Pentazocine has a high affinity for σ1R whereas [3H]DTG bind with equal affinity to both σ1 and σ2 receptors. The σ1R was purified, sequenced and cloned from guinea pig brain in 1996, and bears little sequence homology to any known mammalian receptor5. The crystal structure of the σ1R was reported in 2016, and the reported structure shows it as a trimeric species with a single transmembrane spanning region6. The σ1R is primarily localized in the endoplasmic reticulum (ER), specifically at the mitochondria-associated ER membrane (MAM)7. At the MAM, the σ1R acts as a molecular chaperone for the translocation of inositol-requiring enzyme-1 (IRE-1) in transmitting ER stress signals to the nucleus. The σ1R can also undergo translocation from the ER to the plasma membrane, where it has been found to be associated with a variety of receptors, channels and kinases. This association with a variety of plasma membrane-bound receptor is responsible for the diverse number of biological and pharmacological properties associated with the σ1R.
Although much has been learned about the biology of the σ1R in the two decades since its sequencing and cloning, the same cannot be said of the σ2R. In 2011, Xu et al. reported that the σ2R represented a binding site in a protein complex containing Progesterone Membrane Binding Component-1 (PGRMC1)8. The association of the σ2R with PGRMC1 was initially made via the photoaffinity tagging of this protein with an azido compound having a high affinity for the σ2R. This study did not conclude the σ2R to be PGRMC1, but reported that altering expression levels of PGRMC1 changed the pharmacological properties of small molecules interacting with the σ2R. Evidence supporting this conclusion was reported in two subsequent papers: Izzo et al. reported that knocking down PGRMC1 reduced Aβ oligomer synaptic dysfunction in cultured neurons, and this effect could also be achieved pharmacologically with an antagonist of the σ2R9. Secondly, Yi et al. reported that genetic knockout or knockdown of vem-1, the nematode ortholog of PGRMC1, diminished the neurotoxicity of amyloid precursor protein in a nematode model of AD, and this effect could be demonstrated pharmacologically by using σ2 R antagonists10. Collectively, these data suggested a strong association between the σ2R and PGRMC1. PGRMC1 has been proposed to be a membrane-associated progesterone receptor, but its role in cell biology is also poorly understood11. PGRMC1 is similar to the σ1R in that it too is thought to be a molecular chaperone, in this case one that is involved in the translocation of heme and lipophilic molecules such as cholesterol and other steroids from the plasma membrane and the ER, mitochondria and other organelles12,13. The high association of PGRMC1 and σ2R in breast cancer, and the suspected role of both proteins in cell proliferation, also support an association between these two proteins14,15,16.
Evidence disputing the link between the σ2R and PGRMC1 was first reported by Abate et al., who demonstrated that stable knockdown of PGRMC1 or the overexpression of this protein in MCF7 cells did not alter the binding of [3H]DTG17. A subsequent paper by Chu et al. provided similar results in NSC34 cells where PGRMC1 had been genetically knocked out using CRISPR gene editing techniques18. In each case, the authors concluded that the σ2R was not PGRMC1 and suggested that the data reported by Xu et al. was incorrect17,18. However, the original study did not claim that the σ2R was PGRMC1, only that the σ2R was a binding site located in the complex of proteins that contained PGRMC18. Subsequently, a recent report by Alon et al. concluded that the protein TMEM97 was not only a [3H]DTG binding site, but represented the gene for the σ2R19, raising further doubt about the association between the σ2R and PGRMC120,21.
TMEM97 is a protein that has generated interest lately because of its association with lysosomal cholesterol transporter NPC1, a protein whose loss results in Niemann–Pick disease type C1, a fatal lysosomal storage disorder19. TMEM97 has also been associated with the LDL receptor, and a previous study has shown that siRNA studies knocking down TMEM97 reduced the rate of internalization of LDL by the LDL receptor22. Prior to its identification as TMEM97, the σ2R had also been suggested to be involved in cholesterol biosynthesis13. Consequently, the association of TMEM97/σ2R, the new nomenclature for this protein, with cholesterol pathways was consistent with prior literature reports on the potential role of the σ2R in cell biology. Since PGRMC1 has also been implicated with cholesterol trafficking (reviewed in13), there is a distinct possibility that both TMEM97/σ2R and PGRMC1 may be involved in the same biochemical pathways within the cell. Therefore, as part of our continued interest in the role of the σ2R in cell proliferation, our group conducted a series of CRISPR studies aimed at exploring the functional relationship between TMEM97/σ2R and PGRMC1 on the mechanisms of cholesterol uptake and translocation in the cell. In this report, we present evidence demonstrating that deletion of either TMEM97/σ2R or PGRMC1 reduces the level of internalization and trafficking of LDL by the LDL receptor. Knocking out both proteins in HeLa cells did not have an additive effect on LDL uptake. This effect could also be produced through the pharmacological blocking of TMEM97/σ2R with a σ2R antagonist. These data indicate that both TMEM97/σ2R and PGRMC1 are involved in the internalization of LDL by LDL receptor. Confocal microscopy studies also show that LDLR, PGRMC1 and TMEM97 have an asymmetric distribution in the plasma membrane, suggesting that they are involved in a protein complex. We also report that CRISPR gene editing of TMEM97/σ2R results in a complete reduction of the binding of the σ2R ligand [125I]N-(4-(6,7-dimethoxy-3,4-dihydroisoquinolin-2(1 H)-yl)butyl)-2,3-dimethoxy-5-iodo-benzamide ([125I]RHM-4)23, but only a partial reduction in the specific binding of [3H]DTG to HeLa cell membranes. The residual binding of [3H]DTG was not attributed to binding to the σ1R, and may represent a third, currently uncharacterized sigma receptor.
Results
CRISPR Gene Editing of PGRMC1 and TMEM97
In order to study whether TMEM97 or/and PGRMC1 is responsible for σ2R binding and what their biological functions are, CRISPR technology was used to knockout TMEM97, PGRMC1 and both TMEM97 and PGRMC1 in HeLa cells to generate four knockout (KO) cell lines, i.e. control, TMEM97 KO, PGRMC1 KO, and TMEM97/PGRMC1 double KO. All the four cell lines were generated using the same plasmid transfection and lentivirus-infection procedures. Single clones were isolated for all knockout cell lines and used throughout the current study. Western blot showed that TMEM97 or/and PGRMC1 were completely removed in the knockout cell lines (Fig. 1A). TMEM97 or PGRMC1 protein levels were measured as the expression ratio of TMEM97 or PGRMC1 to GAPDH, respectively, were quantified in control and knockout cell lines. Six clones from each cell line were analyzed. The data showed that TMEM97 KO did not affect PRGMC1 protein level and vice versa (Fig. 1B,C). TMEM97 was knocked out by using three different guide RNA sequences, i.e. sgRNA-1, 2 or 3, each directing cas9 nuclease to cleave distinct, specific site of exon 1 of TMEM97. Western blot analysis showed that TMEM97 proteins were successfully deleted by using each of three sgRNAs (Supplementary Fig. 1). Only sgRNA-1 was used to generate TMEM97 knockout cells for the majority of the studies described below.
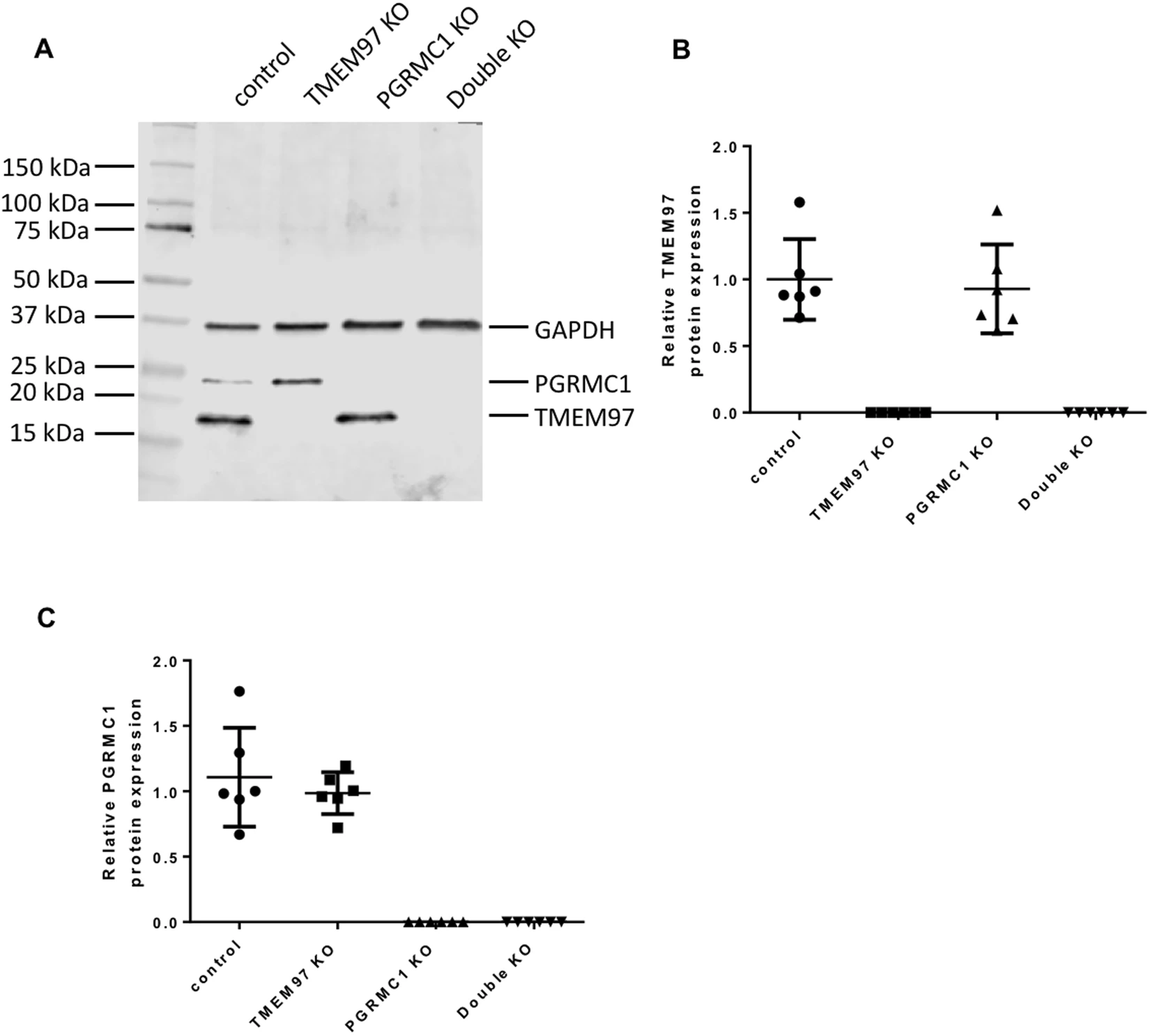
Western blot analysis for control, TMEM97 KO, PGRMC1 KO, and double KO cell lines. All the four cell lines were generated by sequential plasmid transfection and lentivirus-infection procedures. Single clones were isolated for all cell lines. (A) A representative western blot showing that TMEM97 or/and PGRMC1 were completely deleted in the knockout cell lines. (B) TMEM97 and (C) PGRMC1 protein levels measured as the expression ratio of TMEM97 or PGRMC1 to GAPDH, respectively, were quantified in control and knockout cell lines. Six clones from each cell line were analyzed. The data showed that TMEM97 KO did not affect the PRGMC1 protein level and vice versa.
n vitro Binding Studies
In vitro binding studies were conducted using two different radioligands for the σ2R, [125I]RHM-4, which has a high affinity and selectivity for σ2R versus σ1R, and [3H]DTG, which binds equally to σ2R and σ1R. The in vitro binding studies were done in the presence of 1 μM (+)−pentazocine to mask σ1R. The results of the binding studies in the PGRMC1 KO cells indicated that there was no change in the Bmax values for either [125I]RHM-4 and [3H]DTG. However, knocking out PGRMC1 resulted in a small increase in Kd value of both [125I]RHM-4and [3H]DTG (Fig. 2; Table 1). Knocking out TMEM97 resulted in a complete reduction in binding of [125I]RHM-4 to the HeLa cells (Fig. 2B). This effect was observed in the TMEM97 KO cells using all three guide RNA sequences (Supplementary Fig. 2). We also observed a large, but incomplete, reduction of [3H]DTG binding to TMEM97 KO cells (Fig. 2B). The reduction in specific binding of [3H]DTG was observed in TMEM97 KO cells using all three guide RNA sequences, and all cell lines studied retained a residual [3H]DTG binding capacity. To better characterize the residual DTG binding site, saturation ligand binding studies were performed on TMEM97 KO cells using [3H]DTG (Fig. 2C). We determined a Kd value for this residual DTG binding site to be 302 ± 80 nM with a Bmax value of 1218 ± 148 fmol/mg protein.
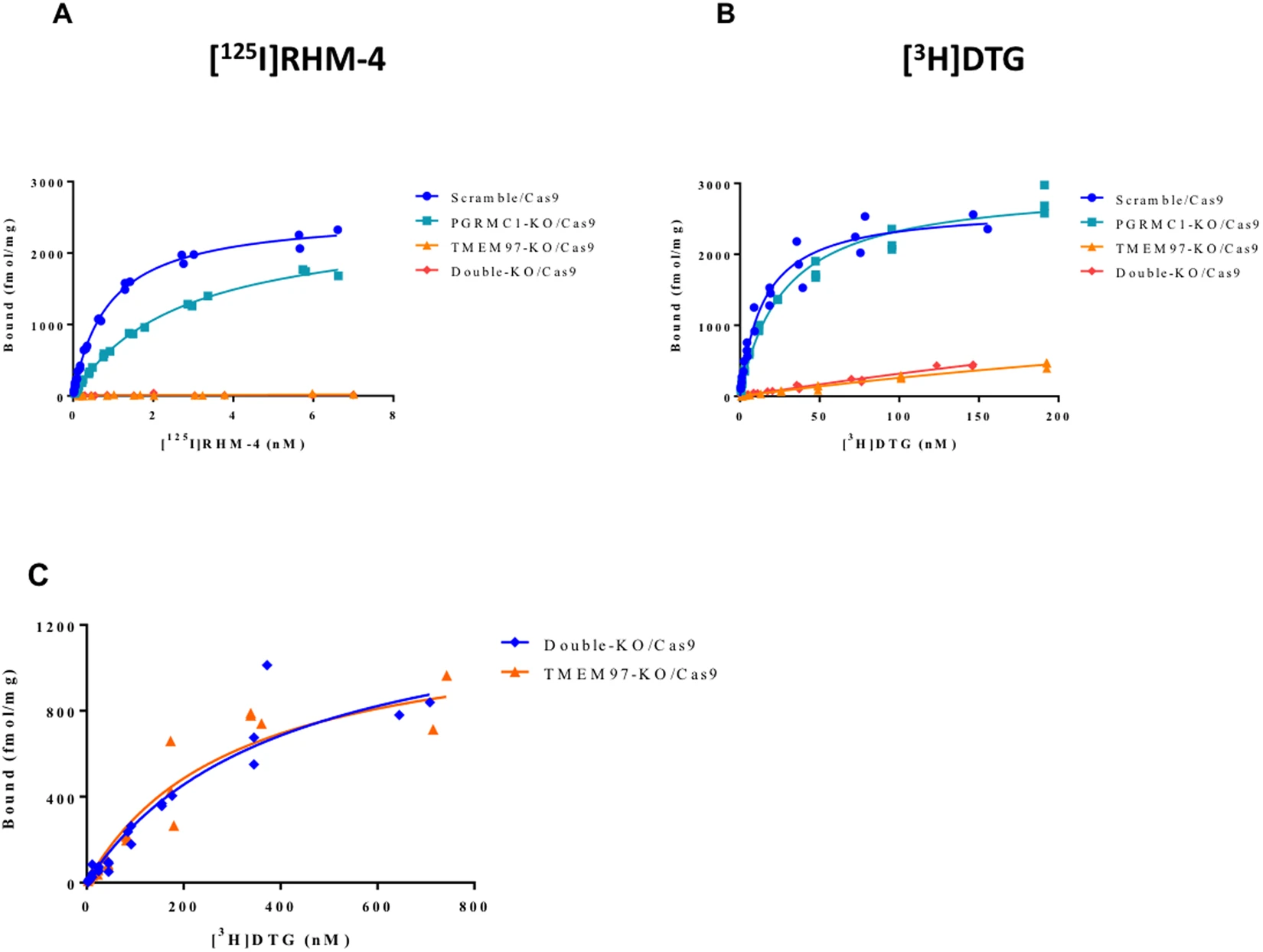
In vitro binding studies in control, TMEM97 KO, PGRMC1 KO, and double KO cell lines. (A) [125I]RHM-4 binding was completely eliminated in the TMEM97 KO and double KO cells. (B) [3H]DTG binding was significantly reduced in the TMEM97 KO and double KO cells, but some residual specific binding was observed. (C) Saturation binding curves were obtained using a ligand concentration of [3H]DTG (1 nM – 750 nM) to obtain Kd and Bmax values for the residual binding site in the TMEM97 KO and double KO cells.
| (KdnM) [3H]DTG | (KdnM) [125I]RHM-4 | Bmax (fmol/mg) [3H]DTG | Bmax (fmol/mg) [125I]RHM-4 | |
|---|---|---|---|---|
| Scramble/Cas9 | 14.8 ± 1.8 | 0.88 ± 0.04 | 2664 ± 105 | 2545 ± 37 |
| PGRMC1 KO/Cas9 | 27.5 ± 2.5 | 2.66 ± 0.12 | 2971 ± 91 | 2475 ± 51 |
| TMEM97 KO/Cas9 | 302.0 ± 80.2 | N.B. | 1218 ± 148 | N.B. |
| Double KO/Cas9 | 402.6 ± 109.8 | N.B. | 1374 ± 192 | N.B. |
In vitro binding data from the radioligand binding studies.
Previous studies have reported that [3H]DTG binds with low affinity to the PGRMC1 homodimer in cells overexpressing this protein24. In order to confirm that this residual binding site did not reflect binding to the PGRMC1 homodimer, in vitro ligand binding studies were conducted in the double KO cells (Fig. 2C). We observed no significant difference in the [3H]DTG Bmax and Kd between the TMEM97 KO and the TMEM97/PGRMC1 double KO cells (Table 1), indicating that the residual binding of [3H]DTG is not related to PGRMC1.
Cancer cells have also been reported to possess splice variants of the σ1R that have low affinity for [3H](+)-pentazocine and [3H]DTG25. In order to confirm that the residual [3H]DTG binding site did not reflect binding to splice variants of the σ1R, Western blots were obtained for the HeLa cells used in this study using an antibody that recognizes all forms of the σ1R. The results of this study revealed that the HeLa cells used in this study contained only the full length σ1R (Data not shown). Therefore, this residual [3H]DTG binding site does not represent binding to a σ1R splice variant.
Interactions between TMEM97, PGRMC1, and the LDL Receptor
Cholesterol, due to its hydrophobic nature, is transported in the bloodstream bound with LDL and is taken up into cells primarily via the binding of LDL to the LDLR followed by receptor-mediated endocytosis26. Since both TMEM97 and PGRMC1 are membrane-bound proteins that have been implicated in cholesterol metabolism, a series of studies were conducted to determine if there is a possible interaction between these proteins and the internalization of LDL by the LDLR. There was no change in protein expression levels of LDLR between the control (Scramble/Cas9), PGRMC1 KO, TMEM97 KO, and double KO cells as assessed by western blot. TMEM97 was not detected in the TMEM97 KO and double KO cell lines, and PGRMC1 was not detected in the PGRMC1 KO cells and double KO cell lines, verifying the knockout of the representative proteins (Fig. 1; Supplementary Fig. 3). Total intracellular cholesterol levels were not affected in the knockout cell lines as assessed by filipin staining (Supplementary Fig. 4).
The next series of experiments studied the effect of knocking out TMEM97 and PGRMC1 on the internalization of LDL by LDLR. Since LDL is internalized via clathrin-mediated endocytosis, we measured the kinetics of LDL internalization using LDL labeled with [3H]cholesterol. Cells were incubated in the presence of lipoprotein-depleted serum (LPDS) for 24 hours in order to induce expression of LDLR. As noted above, there was no difference in LDLR expression between the control cells and any of the knockout cell lines (Supplementary Fig. 3). Uptake of radiolabeled LDL was significantly decreased in PGRMC1 KO, TMEM97 KO, and double KO cell lines. Moreover, this decrease in LDL internalization was not a stepwise reduction with the loss of each protein but rather a complete diminished capability, as PGRMC1 KO and TMEM97 KO cells had the same decreased internalization of radiolabeled LDL as the double KO cells (Fig. 3A). This decreased LDL uptake was also observed in experiments using fluorescently tagged DiI-LDL (Supplementary Fig. 5). To ensure the radiolabeled LDL was taken up via the LDLR pathway, excess unlabeled LDL (250 μg/mL) was incubated with the radiolabeled LDL (50 μg/mL). Under these conditions the excess LDL competes with radiolabeled LDL for binding with LDLR, resulting in significantly decreased internalization (Fig. 3B). These data confirm that the LDL-LDLR pathway is responsible for internalization in these cells.
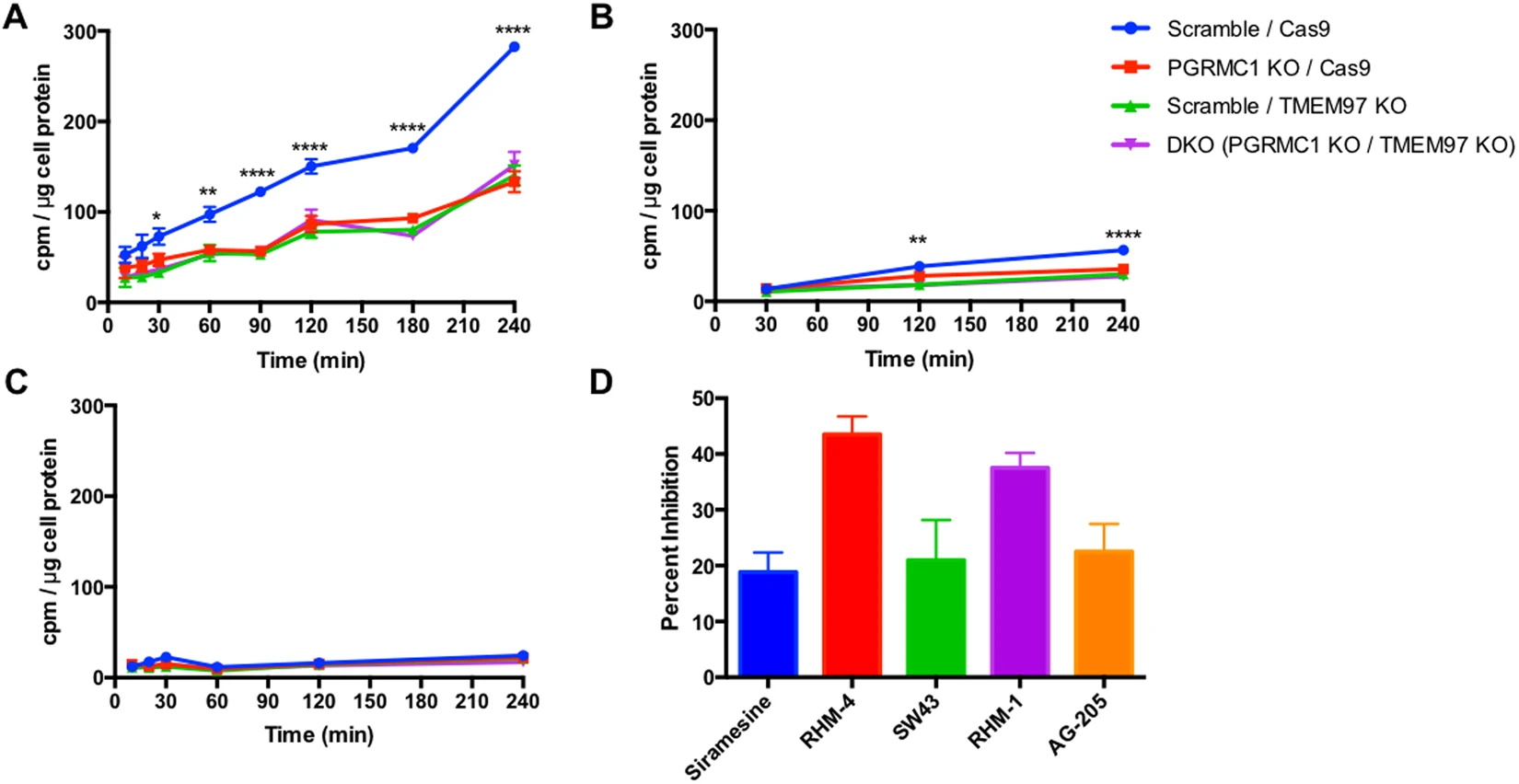
Uptake of [3H]cholesterol labeled LDL (50 μg/mL LDL labeled with 3 μCi/mL [3H]cholesterol) expressed as cpm /μg cell protein. (A) Uptake over time. (B) Uptake over time with the addition of excess unlabeled LDL (250 μg/mL). (C) Incubation at 4 °C to indicate similar capacity of LDL binding to LDL receptor. (D) Percent Inhibition of radiolabeled LDL uptake by incubation with 500 nM Siramesine, RHM-4, SW43, RHM-1, and AG-205. Significance indicated by a 2-way Anova for comparison between cell lines. *Indicates p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, n = 4.
The next set of experiments was conducted to identify if PGRMC1 and TMEM97 are responsible for the initial binding of LDL to the LDLR, or if they are important for the internalization of the bound LDL-LDLR complex. Incubation of HeLa cells in the presence of LDL at 4° allows LDL to bind with LDLR but inhibits receptor-mediated endocytosis of the bound complex27. We observed no significant difference between knockout and control cells in their capacity to bind LDL (Fig. 3C). Their similar capacity to bind LDL further suggests that both PGRMC1 and TMEM97 are involved in the internalization of the LDL-LDLR complex, and they are not involved in the initial binding of LDL to LDLR.
The next logical step was to evaluate the effect of pharmacological inhibition of TMEM97 on the uptake of LDL and compare it to ablation of the protein via knockout. Since TMEM97 is the protein which contains the σ2R, we used two ligands previously described selective σ2R antagonists RHM-4 (Kd ~ 0.2 nM) and RHM-1 (Kd ~ 0.66 nM); two ligands previously described as being σ2R agonists Siramesine (Kd ~ 1.6 nM) and SW43 (Kd ~ 12 nM). We also used AG-205 a PGRMC1 inhibitor (Kd ~ 1 μM against σ2R and Ki~1263 ± 325 to the σ2R for displacing DTG). All compounds resulted in a reduction of LDL uptake, with the classical σ2R antagonists having a greater effect than both the σ2R agonists and the PGRMC1 inhibitor. There was a dose-dependent reduction in uptake of LDL labeled with [3H]cholesterol when HeLa cells were treated with these inhibitors (Supplementary Fig. 6). This pharmacologic inhibition of LDL uptake was observed in control cells only, but did not further decrease uptake in the knockout cells lines, further suggesting that PGRMC1 and TMEM97 must both be present and functional for effective LDL-LDLR complex internalization (Supplementary Fig. 7).
Somatostatin and Insulin Uptake Rates are unaffected in knockout cell lines
To determine whether the presence of TMEM97 and/or PGRMC1 affects all clathrin-mediated endocytic pathways, we examined at the uptake of radiolabeled [125I]TYR11-Somatostatin and [125I] Insulin. Results indicated that there was no difference between the rate of uptake of somatostatin or insulin between the cell lines (Supplementary Fig. 8). This indicates that TMEM97 and PGRMC1 play a unique role in uptake of LDL via their interaction with LDLR.
Confocal Microscopy Studies
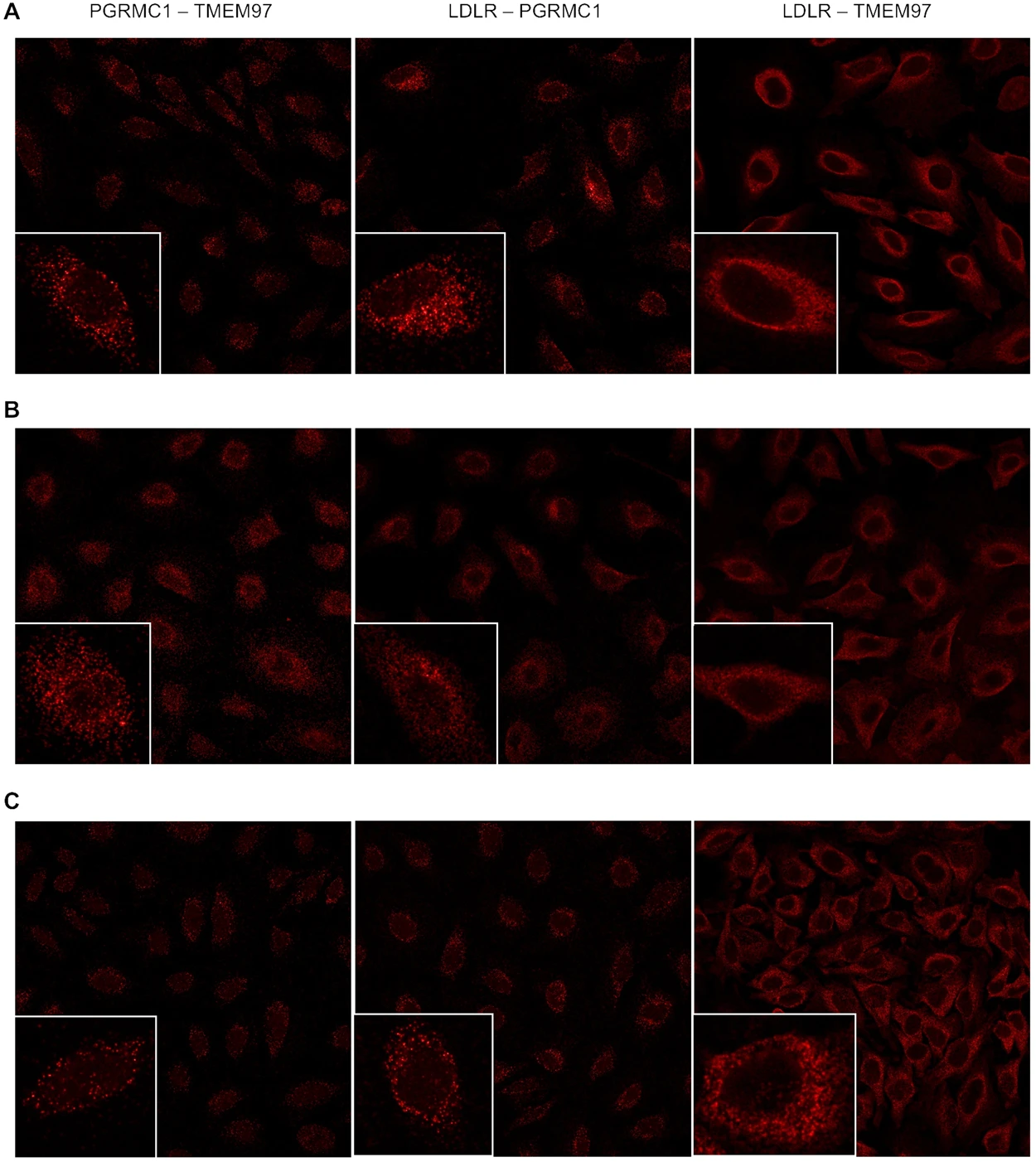
A series of confocal microscopy studies was conducted in order to determine the cellular localization of TMEM97, PGRMC1 and LDLR in HeLa cells. Under serum-starved conditions, there was an asymmetrical distribution of LDLR in the plasma membrane of HeLa cells. This asymmetry of LDLR has been reported previously and is thought to represent localization of LDLR in the region of the plasma membrane rich in lipid rafts. There was a similar asymmetric localization of both TMEM97 and PGRMC1 in the plasma membrane. Co-localization studies indicate that there is a high degree of co-localization of LDLR, TMEM97 and PGRMC1 under these conditions. However, there was a population of PGRMC1 sites that were not co-localized with LDLR and TMEM97 (Fig. 4, Supplementary Fig. 9).
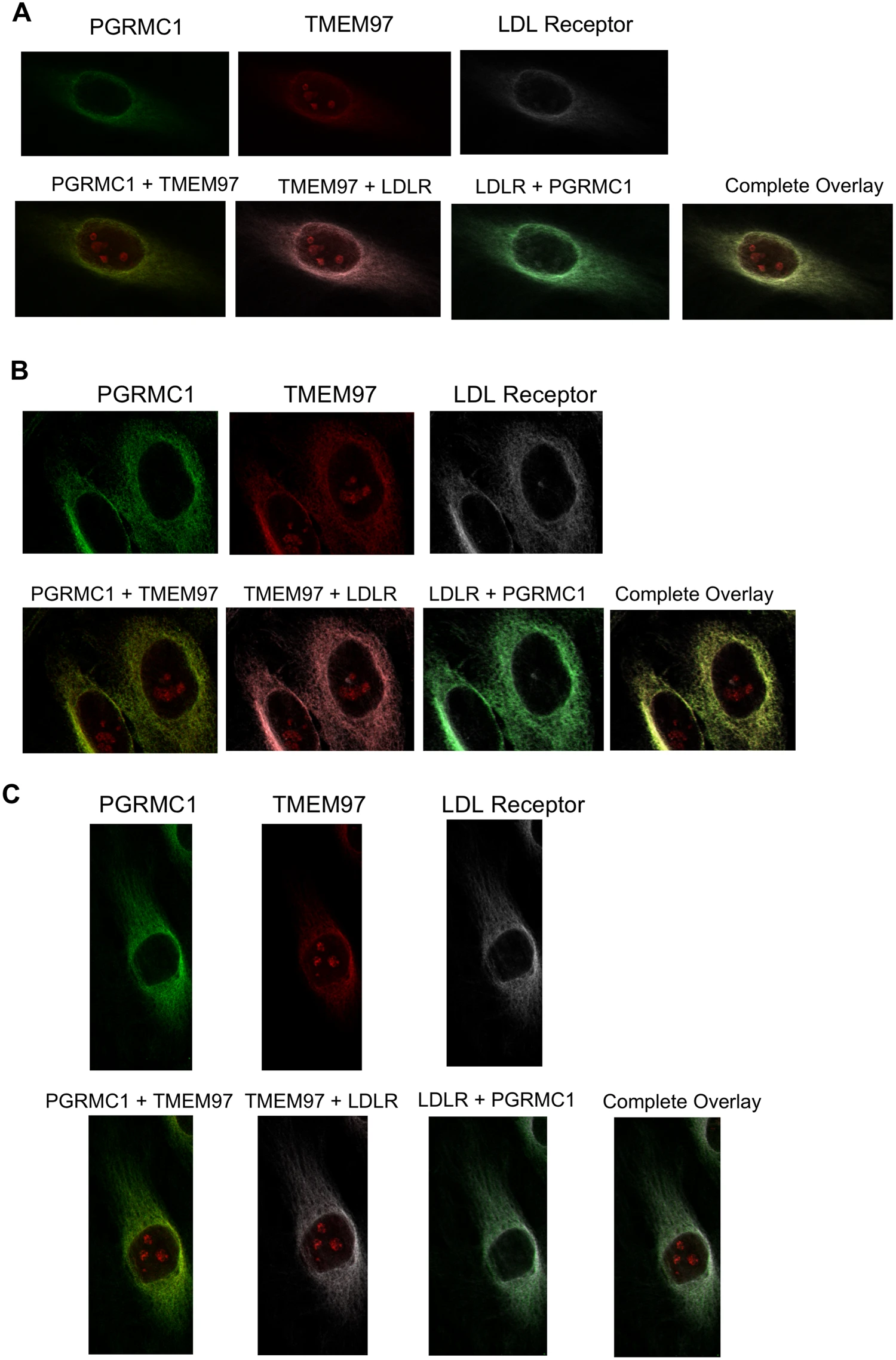
Confocal microscopy on HeLa cell lines labeled with antibodies against LDLR, PGRMC1, and TMEM97. (A) Cells were plated and imaged under standard, non-lipoprotein depleted conditions. (B) Cells were plated and sterol starved by incubation with lipoprotein-depleted serum for 24 hours. (C) Cells were sterol starved for 24 hours, followed by treatment with 50 μg/mL LDL for 3 hours.
The next set of experiments repeated the confocal imaging studies, this time following treatment of previously serum-starved HeLa cells with serum containing LDL. Although there was an asymmetric distribution of LDLR, TMEM97 and PGRMC1 in HeLa cells, and co-localization of the three proteins, there was also a higher percentage of both PGRMC1 and TMEM97 not associated with LDLR. These data are consistent with the dissociation of the LDL-LDLR-TMEM97-PGRMC1 protein complex following internalization into the lysosomal compartments within the cell. The confocal microscopy data, taken collectively with the cell uptake studies of [3H]cholesterol-labeled LDL and fluorescently-tagged LDL, suggest that the rapid internalization of LDL by the LDLR requires the formation of a complex with both PGRMC1 and TMEM97.
Proximity Ligation Assays
To confirm that the observed associations between the protein was indeed a complex formed, proximity ligation assays were conducted pairwise between LDLR, PGRMC1, and TMEM97. We observed interactions between LDLR, TMEM97, and PGRMC1 (Fig. 5) in normal, sterol-starved, and sterol-starved followed by LDL treatment for 3 hours. The interactions between LDLR and TMEM97 and between TMEM97 and PGRMC1 were not observed in the TMEM97 KO and double KO cell lines (Supplementary Fig. 10), as expected. Similarly, the interactions between LDLR and PGRMC1 and between TMEM97 and PGRMC1 were not observed in the PGRMC1 KO and double KO cell lines. However LDLR and PGRMC1 were shown to interact in the TMEM97 KO, and between LDLR and TMEM97 in the PGRMC1 KO, indicating that the LDLR interacts with TMEM97 and PGRMC1 despite the absence of the other protein.
Proximity Ligation Assay on Scramble/Cas9 HeLa cells showing interactions between TMEM97 and PGRMC1, LDLR and PGRMC1, and LDLR and TMEM97. (A) standard, non-lipoprotein depleted conditions, (B) sterol-starved conditions, and (C) sterol-starved conditions followed with treatment with 50 μg/mL LDL for 3 hours. Inserts represent an enlarged region focusing on a single cell.
Taken together this data indicated that the LDLR, TMEM97, and PGRMC1 proteins are associated within close proximity of each other and form a complex.
Discussion
The goal of the current study was to further understand the role of the σ2R in cell biology, particularly its role as a molecular marker of cell proliferation. Previous studies from our group have shown that the density of σ2Rs is higher in proliferating breast cancer cells relative to cancer cells driven into quiescence (i.e., G0) by prolonged serum starvation28,29. PET imaging studies with a σ2R -selective radiotracer, [18F]ISO-1, have shown a strong correlation between radiotracer uptake and the density of σ2R in a murine model of breast cancer30, and an excellent correlation between radiotracer uptake and Ki-67 score in a mixed population of patients diagnosed with lymphoma, breast, and head & neck cancer31. Since the molecular identity of the σ2R was not known at the time of these studies, these encouraging results initiated a series of studies in our lab to characterize this protein as a means of understanding the role of the σ2R in cell proliferation. In 2011, we reported a series of experimental results suggesting that the σ2R was a binding site in a protein complex containing PGRMC18. We did not state that the σ2R was PGRMC1 since we did not observe a 1:1 correlation between the binding of a radioiodinated ligand having a high affinity for the σ2R, and levels of the protein in HeLa cells having either a transient knockdown or overexpression of PGRMC18. Subsequent studies using cancer cells having a stable knockdown or overexpression of PGRMC1 challenged these conclusions since neither condition resulted in a change in binding of the σ2R radioligand, [3H]DTG17,18. The recent identification of the σ2R as being the protein, TMEM97, provided further support to the absence of any correlation between the σ2R, now called the TMEM97/σ2R, and PGRMC119,21.
TMEM97 is thought to be involved in cholesterol trafficking through its association with lysosomal cholesterol transporter NPC1, a protein whose loss results in a fatal lysosomal storage disorder, Niemann–Pick disease type C119. A previous study also demonstrated that siRNA studies knocking down TMEM97 reduced the rate of internalization of LDL by the LDL receptor (LDLR)22. Another protein that has been strongly linked to cholesterol trafficking is PGRMC1, and in a recent review by Cahill and Medlock, a theoretical mechanism showing a role of PGRMC1 in the membrane trafficking of LDLR was proposed32. Therefore, if the σ2R and PGRMC1 are part of a protein complex as we proposed in 2011, the above evidence suggests that this protein complex may consist of the LDLR, TMEM97 and PGRMC1.
In the current study, we provide strong experimental evidence that this is indeed the case. CRISPR gene editing studies resulting in a complete knockout of either TMEM97 or PGRMC1 resulted in an identical reduction in the rate of internalization of 3H-labeled LDL and fluorescently tagged DiI-LDL by LDLR. Knocking out both TMEM97 and PGRMC1 did not lead to a further reduction in 3H-LDL uptake, indicating that the effect was not additive, and loss of one protein has the same effect as losing both proteins. The decreased uptake in the absence of TMEM97, PGRMC1, or both was unique to LDL, as uptake of somatostatin and insulin was not affected, thus demonstrating that knocking out these proteins does not disrupt all clathrin-mediated endocytic pathways. Furthermore, knocking out TMEM97 resulted in a complete loss of binding of [125I]RHM-4, and a significant reduction in the binding of [3H]DTG to HeLa cells. These data indicate that [125I]RHM-4 binds selectively to TMEM97, whereas there is some residual binding of [3H]DTG to a site not associated with TMEM97. The affinity of [3H]DTG for this residual binding site is ~300 nM, and its density is ~40% that of TMEM97. Since this residual binding was also observed in the double KO cells, it does not represent binding to the low affinity DTG binding site identified previously in the PGRMC1 dimer. Whether this site represents a third, previously unidentified sigma receptor or some other off-target binding site remains to be determined. It is also interesting to note that knocking out PGRMC1 resulted in a decrease in affinity of both [125I]RHM-4 and [3H]DTG for TMEM97, but no change in Bmax values for the protein. These results are consistent with an interaction between TMEM97 and PGRMC1, with the knocking out of PGRMC1 resulting in a small change in the 3-dimensional structure of TMEM97 leading to a reduction in affinity of σ2R radioligands binding to this site.
The confocal microscopy and proximity ligation assay studies reported here also support the above conclusions. There was an asymmetric distribution of LDLR, TMEM97, and PGRMC1 in the plasma membrane of HeLa cells under serum-starved conditions33,34. Co-localization studies indicate that there is a high level of co-localization of the three proteins, which is consistent with the formation of an LDLR-TMEM97-PGRMC1 protein complex. However, there was also a population of PGRMC1 apparently not associated with LDLR, as evident in the proximity ligation assay, where the PGRMC1-TMEM97 and LDLR-PGRMC1 pairs showed less colocalization signal than the LDLR-TMEM97 pair. These data are consistent with previous reports that PGRMC1 may be associated with other membrane-bound receptors such as the epidermal growth factor receptor (EGFR) and insulin receptor (IR)22,35. Addition of LDL to HeLa cells after a brief period of serum starvation resulted in the internalization and likely dissociation of the LDL-LDLR-TMEM97-PGRMC1 tetrameric complex in the acidic environment of the lysosomes since this treatment resulted in an increase in the amount of PGRMC1 and TMEM97 not co-localized with LDLR.
The involvement of TMEM97/σ2R in the internalization and trafficking of cholesterol via LDL is consistent with our previous characterization of the σ2R is upregulated in proliferating versus quiescent breast cancer cells. Cholesterol is a key component of the cell membrane and an essential requirement for cell division, and an increase in cholesterol uptake has been demonstrated for proliferating cells36,37,38. Since the demand for cholesterol to support cell proliferation is high, most cancer cells are characterized by an upregulation of the various pathways responsible for its biosynthesis36,39. A disruption of cholesterol biosynthesis has also been proposed as a potential mechanism for treating cancer36,38,39.
The disruption of lipoprotein trafficking may also be responsible for the mechanism of action of σ2R antagonists in preventing Aβ oligomer synaptic dysfunction and Aβ-induced neurotoxicity. Apolipoprotein-E (Apo-E) is the main source of cholesterol in brain, which is required for axonal growth and synaptogenesis. Apo-E is synthesized in astrocytes and microglia and released into the extracellular space where it is taken up in neurons by LDLR and a second lipoprotein receptor, LDL receptor related protein 1 (LRP1). There is a strong correlation between the prevalence of AD and lipoprotein metabolism, in particular apolipoprotein e4 (APO-E4). For example, patients having the APO-E4 allele have a higher incidence of AD and are positive for Aβ plaques as measured by [11C]PiB PET imaging studies at an earlier age. Apolipoproteins also bind Aβ, and it has been proposed that Apo-E-bound Aβ is taken up by neurons via transport by LDLR or LRP1. Therefore, it is possible that σ2R antagonists disrupt the binding of Aβ oligomers or Aβ-bound Apo-E to LDLR or LRP1 in a manner analogous to blocking the uptake of LDL in HeLa cells by the σ2R antagonist, RHM-4.
In summary, our data demonstrate that the rate of uptake of LDL by LDLR is increased by the formation of a complex with both PGRMC1 and TMEM97. Internalization of LDL by LDLR can occur in the absence of this protein complex, but at a much slower rate than what is observed with the ternary complex. Confocal microscopy and radioligand binding studies indicate that PGRMC1 and TMEM97 are physically associated in this complex with LDLR since knocking out PGRMC1 results in a reduction in the affinity of σ2R ligands for TMEM97, and there is a high degree of co-localization of these proteins. The formation of a trimeric LDLR-PGRMC1-TMEM97 complex that results in the rapid internalization and trafficking of LDL is consistent with the conclusion we stated in our 2011 paper, that the σ2R represents a binding site in the protein complex containing PGRMC1.