
The Sigma-2 Receptor / Transmembrane protein 97 (σ2R/TMEM97) Modulator JVW-1034 Reduces Heavy Alcohol Drinking and Associated Pain States in Male Mice

By Sema G. Quadir, Sean M. Tanino, Christian D. Rohl, James J. Sahn, Emily J. Yao, Luíza dos Reis Cruz, Pietro Cottone, Stephen F. Martin, and Valentina Sabino
Excerpt from the article published in Neuropharmacology, Volume 184, 108409, 15 February 2021. DOI: https://doi.org/10.1016/j.neuropharm.2020.108409. Author manuscript available at NCBI HHS public access: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7856185/
Editor’s Highlights
- Feedback between alcohol consumption and pain exists; people suffering from chronic pain self-medicate with alcohol to get relief, which in turn leads to increased pain sensitivity during withdrawal, thus sustaining a negatively reinforced vicious cycle.
- The modulation of σ2R/TMEM97 controls heavy alcohol drinking and blunts the increased hyperalgesia and allodynia that result from excessive alcohol consumption.
- Small molecule modulators of σ2R/TMEM97 have, therefore, the potential to be developed into novel drug candidates for the millions of people suffering from AUD and associated pain states.
Abstract
Alcohol Use Disorder (AUD) is a chronic relapsing disorder characterized by compulsive alcohol intake, loss of control over alcohol intake, and a negative emotional state when access to alcohol is prevented. AUD is also closely tied to pain, as repeated alcohol drinking leads to increased pain sensitivity during withdrawal. The sigma-2 receptor, recently identified as transmembrane protein 97 (σ2R/TMEM97), is an integral membrane protein involved in cholesterol homeostasis and lipid metabolism. Selective σ2R/Tmem97 modulators have been recently shown to relieve mechanical hypersensitivity in animal models of neuropathic pain as well as to attenuate alcohol withdrawal signs in C. elegans and to reduce alcohol drinking in rats, suggesting a potential key role for this protein in alcohol-related behaviors. In this study, we tested the effects of a potent and selective σ2R/TMEM97 ligand, JVW-1034, on heavy alcohol drinking and alcohol-induced heightened pain states in mice using an intermittent access model. Administration of JVW-1034 decreased both ethanol intake and preference for ethanol, without affecting water intake, total fluid intake, or food intake. Notably, this effect was specific for alcohol, as JVW-1034 had no effect on sucrose intake. Furthermore, JVW-1034 reduced both thermal hyperalgesia and mechanical hypersensitivity in ethanol withdrawn mice. Our data provide important evidence that modulation of σ2R/TMEM97 with small molecules can mediate heavy alcohol drinking as well as chronic alcohol-induced heightened pain sensitivity, thereby identifying a promising novel pharmacological target for AUD and associated pain states.
1. Introduction
Alcoholism represents a serious global public health problem. In the United States, the number of alcohol-related deaths has nearly doubled between 1999 and 2017 (White et al., 2020), and the total cost of alcohol problems is estimated at $249 billion a year (Sacks et al., 2015). According to the 2018 NSDUH, in the United States 14.4 million adults ages 18 and older (5.8%) had alcohol use disorders (AUD) (2020).
AUD is a chronic relapsing disorder characterized by compulsive alcohol intake, loss of control over consumption, and a negative emotional state when access to alcohol is prevented (Becker and Koob, 2016; Koob, 2013). An exacerbating feedback between alcohol consumption and pain exists; people suffering from chronic pain self-medicate with alcohol to get relief (Angst and Clark, 2006; Brennan et al., 2005; Gatch, 2009; Riley and King, 2009), which in turn leads to increased pain sensitivity during withdrawal, thus sustaining a negatively reinforced vicious cycle (Avegno et al., 2018; Egli et al., 2012; Koob, 2008). Pain hypersensitivity in chronic alcohol drinkers arises from the emergence of alcoholic neuropathy and manifests as both allodynia (when a non-noxious stimulus induces a nociceptive response) and hyperalgesia (when a noxious stimulus produces a heightened nociceptive response) (Arout et al., 2016; Egli et al., 2012; Koike et al., 2001). Even though pain is most commonly regarded as a peripheral pathology, a central sensitization of ascending nociceptive signaling is evident in chronic pain states including those produced by chronic alcohol (Apkarian et al., 2013; Latremoliere and Woolf, 2009; Viswanath et al., 2020; Xu et al., 2020), and neurocircuitries of AUD and neuropathic pain have been found to overlap substantially (Egli et al., 2012; Robins et al., 2019). Therefore, therapeutic strategies that could successfully treat both excessive drinking and heightened pain sensitivity in individuals with AUD could be of particular value.
The two subtypes of sigma receptors (σRs) have been proposed to be promising therapeutic targets for AUD (Schmidt and Kruse, 2019; Smith, 2017). The sigma-1 receptor (σ1R) is a unique pharmacologically regulated molecular chaperone that serves as a scaffolding protein and modulates the activity of several associated proteins (Hayashi and Su, 2007; Oyer et al., 2019). What had historically been referred to as the sigma-2 receptor (σ2R) has recently been identified as transmembrane protein 97 (TMEM97), which is an integral membrane protein involved in cholesterol homeostasis and lipid metabolism (Alon et al., 2017; Bartz et al., 2009; Ebrahimi-Fakhari et al., 2016; Matsumoto, 2009; Wilcox et al., 2007). The identification of σ2R as TMEM97, which will be referred to herein as σ2R/TMEM97 for the murine protein that is relevant to this study, will greatly facilitate efforts aimed at probing its function in cellular and physiological processes and disease. σ2R/TMEM97 has long been referred to as a receptor, although no endogenous ligand has been definitively identified, and no intrinsic enzymatic or signaling activity is known. Nevertheless, terms such as agonist and antagonist have been commonly used in the literature, but because reliable functional assays for σ2R/TMEM97 are not available (Oyer et al., 2019; Zeng et al., 2019), these descriptors may not be appropriate. Hence, compounds that target σ2R/TMEM97 will be referred to herein as “modulators.”
The role of σ1R in various disease states including alcohol addiction has been investigated for a number of years (Cottone et al., 2012; Quadir et al., 2019; Sabino et al., 2011; Sabino et al., 2009a; Sabino et al., 2009b). However, because σ2R/TMEM97 had long been only a pharmacologically defined entity, progress toward exploring the effects of engaging this target have been slow, although it has been implicated in cancer (Abate et al., 2018; Baiamonte et al., 2014; Huang et al., 2014). This situation began to change several years ago when it was shown that small molecules that are known to bind selectively to this receptor have neuroprotective properties and beneficial effects in transgenic animal models of Alzheimer’s disease (Izzo et al., 2014a; Izzo et al., 2014b; Yi et al., 2017). Indeed, compounds with high affinity for σ2R/TMEM97 are currently being tested in clinical trials for imaging in breast cancer as well as for treatment of Alzheimer’s disease and schizophrenia (Clinical-Trial-NCT02284919, 2019; Clinical-Trial-NCT02907567, 2018; Clinical-Trial-NCT03397134, 2020). Several compounds that bind to σ2R/TMEM97 have recently been shown to relieve mechanical sensitivity in the spared nerve injury model of neuropathic pain (Sahn et al., 2017). In another study, a σ2R/TMEM97 modulator was found to improve cognitive performance and reduce axonal degeneration in a blast model of traumatic brain injury (TBI) as well as to improve survival of cortical neurons and oligodendrocytes in controlled cortical impact injury model of TBI (Vazquez-Rosa et al., 2019). As a prelude to the present study, we have recently shown that JVW-1034, a small molecule modulator of σ2R/TMEM97, attenuates alcohol withdrawal signs in C. elegans specifically via σ2R/TMEM97 and reduces alcohol drinking in ethanol-dependent rats, suggesting a potential key role for this protein in alcohol-related behaviors (Scott et al., 2018). Here, we now investigated the effect of JVW-1034 on both heavy alcohol drinking and alcohol-induced heightened pain states in mice using a chronic, intermittent access to alcohol model.
…
3. Results
3.1. Effect of JVW-1034 on Ethanol Intake
We found a statistically significant effect of JVW-1034, the structure of which is shown in Fig. 2A, on ethanol intake [Dose: F(2,16)=6.08, p< 0.05; Time x Dose: F(2,16)=0.78, n.s.]; post-hoc analysis showed that the 30 mg/kg dose reduced alcohol intake by 55% at the 2 h time point and 38% after 6 h, as shown in Fig. 2B. Water intake was not significantly affected by JVW-1034 (Fig. 2C) [Dose: F(2,16)=1.68, n.s.; Time x Dose: F(2,16)=1.05, n.s.]. Preference for ethanol was significantly affected by JVW-1034 [Dose: F(2,16)=7.45, p<0.001; Time x Dose: F(2,16)=2.28, n.s.], as shown in Fig. 2D; post-hoc analysis showed that the 30 mg/kg dose decreased preference by 49% at 2 h and 30% at the 6 h timepoints. No effect of JVW-1034 on fluid intake was observed [Dose: F(2,16)=0.14, n.s.; Time x Dose: F(2,16)=0.63, n.s. ] (not shown) nor on food intake [Dose: F(2,16)=2.08, n.s.; Time x Dose: F(2,16)=2.23, n.s.], as shown in Fig. 2E. The effects of drug on ethanol intake and preference did not last until the 24 h time point [ethanol intake, Dose: F(2,16)=1.67, n.s.; Preference, Dose: F(2,16)=0.37, data not shown].
3.2. Effect of JVW-1034 on Sucrose Intake
We found no effect of JVW-1034 on sucrose intake [Dose: F(2,12)=0.45, n.s.; Time x Dose: F(2,12)=1.26, n.s.] (Fig. 3A). We found an effect of JVW-1034 on water intake [Time x Dose: F(2,12)=1.56 n.s.; Dose: F(2,12)=4.96, p<0.05] with 10 mg/kg JVW-1034 increasing water intake by 270% at the 2 h timepoint and 150% at the 6 h timepoint (Fig. 3B). There was no effect at 24 h on sucrose intake, while the effect on water persisted [Sucrose, dose: F(2,12)=1.31, n.s.; Water, dose: F(2,12)=12.6, p<0.05, data not shown].
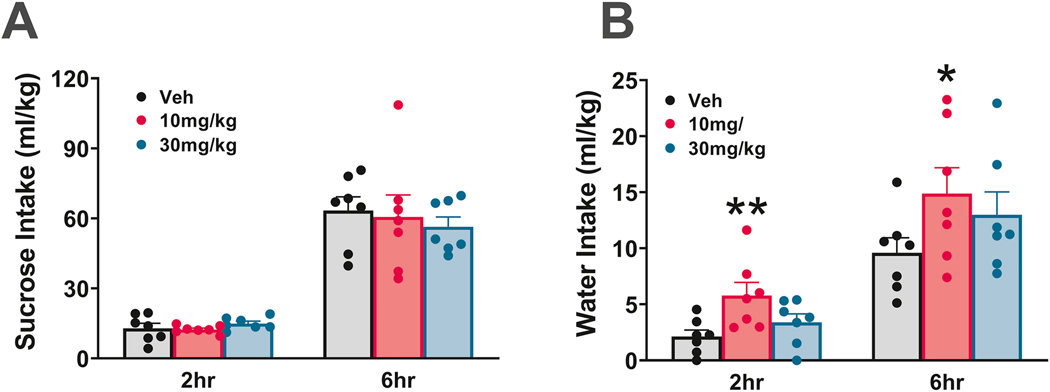
Effect of JVW-1034 on (A) sucrose intake and (B) water intake. N=7. Data represent Mean ± SEM. * p< 0.05, ** p< 0.01 vs. Veh (Newman Keul’s test).
3.3. Effect of JVW-1034 on Mechanical Pain Sensitivity
Alcohol experienced mice showed higher mechanical pain sensitivity in the von Frey test during withdrawal, compared to control mice (Ethanol: F(1,10)=9.53, p=0.011). We also found a significant effect of JVW-1034 on mechanical pain sensitivity [Dose x Ethanol x Force: F(2,20)=6.67 p<0.01; Dose x Ethanol: F(1,10)=37.67 p<0.001; Dose: F(1,10)=25.03 p<0.001], as shown in Fig. 4A. At 72 h withdrawal, a 10%, 35% and 28% reduction in paw withdrawal force (at applied forces of 4 g, 6 g and 8 g respectively) was observed in vehicle-treated mice, which was fully reversed by the administration of JVW-1034 30 mg/kg (the dose effective on alcohol intake). On the other hand, JVW-1034 did not affect mechanical pain sensitivity in water-drinking mice, indicating the effect is specific to the ethanol-drinking mice.
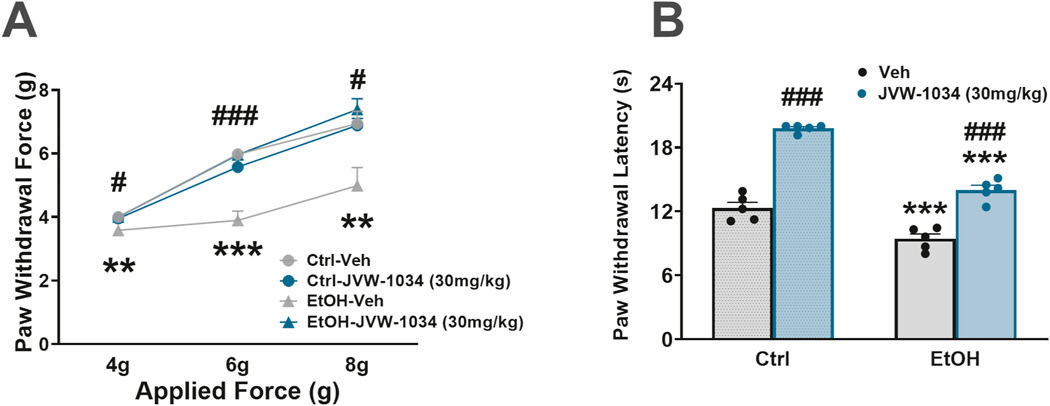
Effect of JVW-1034 on (A) mechanical sensitivity and (B) thermal sensitivity. N=6/group. Data represent Mean ± SEM. ** p< 0.01, *** p< 0.001 vs. Veh; ### p< 0.001 vs. Ctrl (Newman Keul’s test).
3.4. Effect of JVW-1034 on Thermal Pain Sensitivity
Alcohol experienced mice showed higher thermal pain sensitivity in the Hargreaves test during withdrawal, compared to control mice (Ethanol: F(1,16)=102.88, p<0.001). JVW-1034 significantly affected paw withdrawal latency in this test [Dose: F(1,16)=197, p<0.001; Dose x Ethanol: F(1,16)=11.40, p<0.01], as shown in Fig. 4B. EtOH-Veh mice showed, 24 h into withdrawal, a 24% decrease in latency to paw withdrawal compared to Ctrl-Veh mice. When JVW-1034 30 mg/kg (the dose effective on alcohol intake) was administered 30 min prior to the test, this resulted in a 49% increase in paw withdrawal latency in EtOH-JVW-1034 mice, compared to EtOH-Veh mice. Interestingly, when administered to Control, ethanol-naïve mice, JVW-1034 also increased paw withdrawal latency by 61% compared to Ctrl-Veh mice.
3.5. Effect of JVW-1034 on Locomotor Activity
We found no effect of JVW-1034 on locomotor activity across the 10-min bins [Dose: F(1,16)=0.62, n.s.; Dose x Ethanol x Time: F(11,176)=1.00, n.s.; Dose x Ethanol: F(1,16)=0.00, n.s.] (Fig. 5A). We also saw no effect when examining total beam breaks during the entire 2h period of observation [Dose: F(1,16)=0.62,n.s.; Dose x Ethanol: F(1,16)=0.00, n.s.], as shown in Fig. 5B.
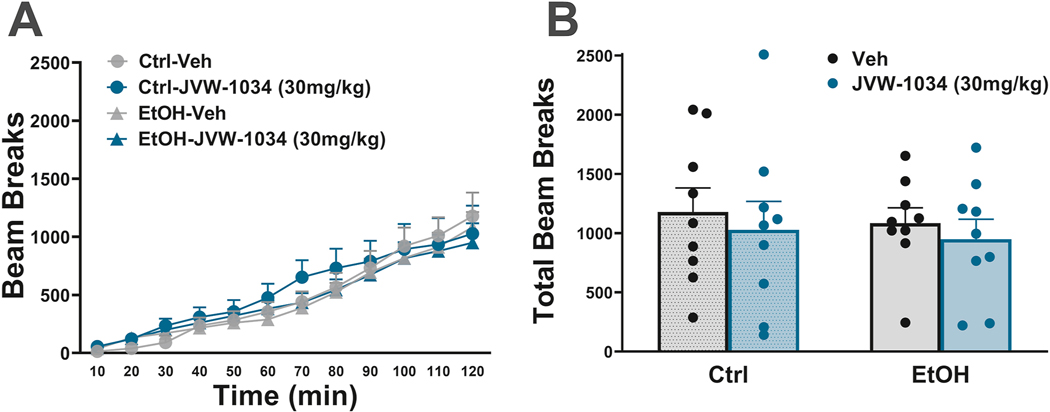
Effect of JVW-1034 on locomotor activity (A) across time and (B) total. N=9 per group. Data represent Mean ± SEM.
4. Discussion
We have previously shown that the σ2R/TMEM97 modulator, JVW-1034, improves behavioral impairments in C. elegans that were withdrawn from chronic exposure to ethanol (Scott et al., 2018), and the activity of this compound was shown to depend upon both the TMEM97 ortholog in the worm as well as its partner protein progesterone receptor membrane component 1 (PGRMC1) (Riad et al., 2018). Furthermore, JVW-1034 significantly reduced voluntary ethanol intake in rats made dependent via the exposure to chronic ethanol vapors, but it did not affect water intake in dependent animals or the lower ethanol intake of non-dependent animals (Sahn et al., 2017; Scott et al., 2018). These pioneering discoveries showed that small molecules that selectively bind to σ2R/TMEM97 can mitigate behaviors resulting from chronic exposure to alcohol. In an extension of those investigations, we utilized JVW-1034 in the present series of experiments to probe the role of σ2R/TMEM97 in excessive alcohol drinking and associated pain states using an intermittent, two bottle choice access to ethanol mouse model. We found that JVW-1034 decreased both ethanol intake and preference without affecting water, total fluid or food intake. Notably, this effect was specific for alcohol, as JVW-1034 had no effect on sucrose intake. JVW-1034 also reduced both thermal and mechanical pain hypersensitivity caused by ethanol withdrawal.
Heavy drinking can be modeled by exposing rodents to repeated cycles of voluntary access to ethanol followed by withdrawal. The model used here is the intermittent access two bottle choice originally proposed by Wise (Wise, 1973), which yields high levels of ethanol intake, as well as behavioral, neurochemical, and molecular adaptations (Bloodgood et al., 2020; Carnicella et al., 2008; Carnicella et al., 2014; Newman et al., 2018; Simms et al., 2008; Zhou et al., 2017). We observed that mice treated with JVW-1034 showed a selective reduction of alcohol intake. These results are consistent with our prior work in which we found promising anti-ethanol effects of this σ2R/TMEM97 modulator using a different species (Scott et al., 2018). The current observations that JVW-1034 reduces voluntary ethanol in a mouse model of heavy drinking imply that the findings are generalizable to multiple species, thus supporting our hypothesis that σ2R/TMEM97 represents a new and highly promising therapeutic target for AUD. The reduction of ethanol intake is selective, as sucrose solution intake or concurrent food or water intake are not affected, and there is no reduction of motor activity. These notable findings suggest that JVW-1034 does not produce malaise-like, sedative, or other nonspecific behavior-impairing effects.
Several lines of pharmacological evidence have suggested a possible role for σ2R/TMEM97 in addiction. For example, (±)-SM 21 and SN79, which are known compounds with high and preferential affinity for σ2R/TMEM97 relative to the σ1R subtype, attenuate behavioral as well as toxic effects of cocaine (Kaushal et al., 2011; Lever et al., 2014; Matsumoto and Mack, 2001; Matsumoto et al., 2007; Mesangeau et al., 2008). The highly selective σ2R/TMEM97 ligand siramesine (Lu-28–179) also reverses the behavioral and molecular effects of cocaine (Kaushal et al., 2011; Klawonn et al., 2017; Matsumoto et al., 2007; Nuwayhid and Werling, 2006).
Even though our study employed systemic injection of JVW-1034, we hypothesize the actions are occurring centrally, and there are several studies that suggest modulation of dopaminergic systems might be involved. For example, within-system neuroadaptations occur in the dopaminergic system as a consequence of chronic alcohol drinking, and these changes may result in a hypodopaminergic state (Koob, 2013). Activation of σ2R/TMEM97 has been shown to lead to dopamine release in the nucleus accumbens (Garces-Ramirez et al., 2011), and rats trained to lever-press for cocaine will maintain lever pressing rates when cocaine is substituted with the σ1R- and σ2R/TMEM97-binding ligand ditolylguanidine (DTG) (Hiranita et al., 2010). An alternative mechanism of action might involve σ2R/TMEM97-mediated inhibition of dopamine D1signaling, since σ2R/TMEM97 activation has been shown to attenuate D1 agonist-induced increases in cAMP levels via a physical interaction (Aguinaga et al., 2018).
Increases in both mechanical and thermal sensitivity have been shown in alcohol withdrawal (Avegno et al., 2018; Dina et al., 2006; Edwards et al., 2012; Robins et al., 2019; Roltsch Hellard et al., 2017; Smith et al., 2017). Medications currently available for AUD focus on reducing alcohol intake and/or relapse to it, but they rarely target the negative states present during alcohol withdrawal, which include intense pain states (Anton et al., 2006; Garbutt, 2009; Garbutt et al., 1999). Hence, therapeutic strategies that successfully treat not only the excessive drinking but also the heightened pain sensitivity that arises during alcohol withdrawal are of particular value (Egli et al., 2012), whether they act through the same or different mechanisms (e.g., central vs. peripheral or same vs. different brain regions). Interesting follow-up studies could determine whether pretreatment with σ2R/TMEM97 modulators are able to counteract other aversive states associated with the absence of alcohol, such as affective signs of withdrawal.
Our finding of the anti-hyperalgesic and anti-allodynic effects of σ2R/TMEM97 modulation in alcohol withdrawal is consistent with a previous study in non-alcohol pain models. For example, the σ2R/TMEM97 modulators siramesine, DKR-1005, DKR-1051 and UKH-1114 reduce mechanical sensitivity in a spared nerve injury model of neuropathic pain (Sahn et al., 2017). Interestingly, neither DKR-1051 nor UKH-1114 improves behavioral impairments in C. elegans that have been withdrawn from chronic exposure to ethanol (Scott et al., 2018). These collective observations suggest that the anti-alcohol behavior induced by JVW-1034 and the anti-pain effects produced by DKR-1051 and UKH-1114 may occur following the binding of these compounds to σ2R/TMEM97 in distinct poses, thereby resulting in the activation of different downstream pathways.
The specific site of the analgesic and anti-neuropathic effects of small molecule interactions with σ2R/TMEM97 may be the spinal cord or the dorsal root ganglion (DRG), where σ2R/TMEM97 is heavily expressed and where σ2R/TMEM97 may modulate the N-methyl-D-aspartate receptor and protein kinase C epsilon activation (Dina et al., 2000; Roh et al., 2010; Sahn et al., 2017). However, a central mechanism of action cannot be ruled out, considering that central sensitization of ascending nociceptive signaling is evident in chronic pain states, including those produced by chronic alcohol exposure (Apkarian et al., 2013; Latremoliere and Woolf, 2009; Viswanath et al., 2020; Xu et al., 2020). In particular, σ2R/TMEM97 is highly expressed in the anterior cingulate cortex (ACC), one of the cortical areas most frequently linked to the emotional reaction to pain and whose hyperexcitability results in pain-induced aversion (Bouchard and Quirion, 1997; Lieberman and Eisenberger, 2015; Meda et al., 2019; Moisset and Bouhassira, 2007; Price, 2000; Sahn et al., 2017). Thus, JVW-1034 may exert its antinociceptive effects by modulating σ2R/TMEM97 in the ACC. Interestingly, while the effect of JVW-1034 on mechanical sensitivity was selective for alcohol experienced mice because it normalized the lower pain threshold in alcohol withdrawn mice but did not affect it in controls. JVW-1034 also affected thermal sensitivity in ethanol-naïve mice. This observation suggests that while σ2R/TMEM97 may be tonically involved in the transmission of thermal pain, it only plays a role in mechanical pain when allodynia develops.
These studies coupled with our previous report (Scott et al., 2018) show that JVW-1034 helps control heavy alcohol drinking and reduces chronic alcohol-induced pain sensitivity. Although little is known about the function of σ2R/TMEM97, we can speculate that these effects arise because binding of JVW-1034 to σ2R/TMEM97 selectively modulates the interaction of σ2R/TMEM97 with other proteins. Indeed, σ2R/TMEM97 is known to associate with a number of membrane proteins, including PGRMC1 and the low density lipoprotein receptor (Riad et al., 2018), Nieman-Pick C1 protein (NPC1) (Bartz et al., 2009; Ebrahimi-Fakhari et al., 2016) as well as σ1R and the D1 receptor (Aguinaga et al., 2018). In addition, we have shown that structurally similar, yet distinct, compounds known to bind selectively to σ2R/TMEM97 can have different, sometimes opposing, effects in animal and cell-based models of neurodegenerative and neurological conditions (Yi et al., 2017; Sahn et al., 2017; Scott et al., 2018). Based upon these observations, we can hypothesize that structurally distinct ligands interact differently with the putative binding site of σ2R/TMEM97, thereby stabilizing one of multiple active conformational states that is in turn specifically recognized by other associated proteins. Variations in the specificity and nature of these protein-protein interactions then lead to defined downstream activities and outcomes. Other mechanisms of action are of course possible and further studies will be needed to elucidate how modulation of σ2R/TMEM97 with JVW-1034 influences alcohol-induced effects.
One limitation of our study is that it was only performed in male mice; to date, there is no evidence suggesting sex differences with respect to σ2R/TMEM97 receptor expression or response to its modulators, but future studies will be needed to address this in the context of alcohol drinking and pain sensitivity. In addition, this study did not measure blood alcohol levels in mice treated with JVW-1034 and, therefore, potential effects on the pharmacokinetics of alcohol cannot be completely ruled out.
5. Conclusions
In summary, the results of the present study provide persuasive evidence that modulation of σ2R/TMEM97 controls heavy alcohol drinking and blunts the increased hyperalgesia and allodynia that result from excessive alcohol consumption. These findings are highly significant; the σ2R/TMEM97-mediated pathway is becoming increasingly associated with the effects of alcohol and small molecule modulators of σ2R/TMEM97 have, therefore, the potential to be developed into novel drug candidates for the millions of people suffering from AUD and associated pain states.