
Negative Impact of Sigma-1 Receptor Agonist Treatment on Tissue Integrity and Motor Function Following Spinal Cord Injury
By Alise Lattard, Gaëtan Poulen, Sylvain Bartolami, Yannick N. Gerber, and Florence E. Perrin
Excerpt from the article published in Frontiers in Pharmacology, 10 February 2021
Sec. Neuropharmacology, Volume 12 – 2021, DOI: https://doi.org/10.3389/fphar.2021.614949
Editors Highlights
- Mitochondrial dysfunction is an essential component of the secondary injury cascade induced by Traumatic spinal cord injury (SCI).
- Sigma-1 receptor agonists favor brain plasticity and induce protective effect in the central nervous system (CNS) under pathological conditions but on the other hand isolated mouse brain mitochondria exposed to σ1R agonists display an increase in reactive oxygen species (ROS) level.
- The present study therefore suggests the detrimental effect of σ1R agonist on motor recovery, neuronal preservation and astrogliosis following SCI.
Abstract
In traumatic spinal cord injury, the initial trauma is followed by a cascade of impairments, including excitotoxicity and calcium overload, which ultimately induces secondary damages. The sigma-1 receptor is widely expressed in the central nervous system and is acknowledged to play a key role in calcium homeostasis. Treatments with agonists of the sigma-1 receptor induce beneficial effects in several animal models of neurological diseases. In traumatic injury the use of an antagonist of the sigma-1 receptor reversed several symptoms of central neuropathic pain. Here, we investigated whether sigma-1 receptor activation with PRE-084 is beneficial or detrimental following SCI in mice. First, we report that PRE-084 treatment after injury does not improve motor function recovery. Second, using ex vivo diffusion weighted magnetic resonance imaging completed by histological analysis, we highlight that σ1R agonist treatment after SCI does not limit lesion size. Finally, PRE-084 treatment following SCI decreases NeuN expression and increases astrocytic reactivity. Our findings suggest that activation of sigma-1 receptor after traumatic spinal cord injury is detrimental on tissue preservation and motor function recovery in mice.
Introduction
Traumatic spinal cord injury (SCI) results in 0.6–0.9 million annual new cases worldwide (Kumar et al., 2018). SCI symptoms include sensory, motor and autonomic deficits ranging from minimal dysfunctions to complete tetraplegia. Following the primary traumatic mechanical disruption of spinal cord tissues, a cascade of events including vascular impairment, mitochondrial dysfunction, excitotoxicity and calcium overload characterizes the secondary phase of injury and further exacerbates the lesion. There is currently no curative treatment to improve neurological recovery after SCI.
The sigma-1 (σ1) receptor is a unique non-G-protein-coupled membrane-associated protein ubiquitously expressed with chaperone activity (Hayashi and Su, 2007). The central nervous system (CNS) is one of the major sites of σ1R activity. Indeed, σ1R is expressed in neurons, astrocytes, oligodendrocytes and microglia and is found at the endoplasmic reticulum (ER) membrane that is associated with the mitochondria (Mitochondria-Associated ER Membranes [MAMs]) [for review see (Penke et al., 2018)]. σ1R, that forms a complex at MAMs with BiP (binding immunoglobulin protein), another chaperone protein, is a Ca2+ sensitive and ligand-operated receptor chaperone at MAMs (Hayashi and Su, 2007). In fact, σ1R modulates Ca2+ exchange between ER and mitochondria by interacting with inositol-1,4,5 triphosphate receptors (IP3Rs) (Hayashi and Su, 2007). Following ligand stimulation, σ1R dissociates from BiP, conducting to a lengthened Ca2+ signaling into mitochondria. Decreasing σ1R activity in cell enhances apoptosis, whereas increasing it neutralizes ER stress response (Hayashi and Su, 2007). σ1R is amongst others implicated in neuroplasticity, neuroprotection and carcinogenesis (Hayashi and Su, 2007). Interestingly, newborn σ1R knockout mice, display a transient enhanced proliferation of progenitor cells in the hippocampal dentate gyrus followed by a decrease in survival and neurite outgrowth of newly generated neurons associated with a reduced function of N-methyl-D-aspartate receptor (NMDAr) (Sha et al., 2013).
PRE-084 (2-(4-morpholinoethyl)-1-phenylcyclohexane-1-carboxylate hydrochloride) is a highly selective σ1R agonist displaying minimal cross reactivity with other receptors [for extensive review see (Motawe et al., 2020)]. It had been shown in vivo that PRE-084 binds with σ1R and is rapidly distributed in the CNS (Motawe et al., 2020). Using excitotoxic injury in organotypic spinal cord slices, Guzmán-Lenis et al. demonstrated that PRE-084 not only induces a neuroprotective effect and decreases neuronal damage but also enhances axonal re-growth (Guzman-Lenis et al., 2009). Furthermore, σ1R activation attenuates several aspects of microglial activation in primary culture of rat microglia (Hall et al., 2009).
In vivo, the beneficial effects of PRE-084 has been reported in animal models of neurological diseases such as Alzheimer disease, Parkinson disease, amyotrophic lateral sclerosis, spinal muscular atrophy, Huntington disease, vascular dementia, amnesia, embolic stroke and multiple sclerosis. Though, PRE-084 did not improve outcomes following seizures [for review see (Motawe et al., 2020)].
To our knowledge, PRE-084 impact has not been investigate following acute traumatic injury in the CNS. However, it had been recently shown that after spinal cord contusion in mice, treatment with a σ1R antagonist reduced mechanical allodynia and thermal hyperalgesia that are characteristics of central neuropathic pain (Castany et al., 2018; Castany et al., 2019). On the one hand, treatment with σ1R agonist plays beneficial roles in chronic neurological diseases, including amelioration of motor-induced symptoms. On the other hand, treatment with σ1R antagonist induce positive outcomes on injury-induced pain following acute CNS injury. It is therefore of interest to investigate the role of σ1R activation on motor function recovery following SCI.
In this study, we assessed the effect of PRE-084 treatment following lateral hemisection of the mouse spinal cord on motor recovery and spinal cord structure at tissue and cellular levels.
Results
σ1R Agonist Treatment After Spinal Cord Injury Does Not Improve Motor Function and Preservation of Neuromuscular Junction Density
To investigate whether activation of σ1receptor after SCI is beneficial to motor recovery, we administered an oral 6-weeks PRE-084-treatment (1 mg/kg/day) to mice starting immediately after thoracic nine lateral hemisection of the spinal cord. As a prerequisite we verified that all behavioral parameters were affected by the injury (Figures 1A–H). PRE-084 did not modify the weight curves over the course of the experiment neither in uninjured nor injured mice (Figure 1A). We studied locomotion of the animal over the treatment period. PRE-084 did not improve the general spontaneous motricity analyzed by open field in both uninjured and injured groups (Figure 1B). The regularity index that reflects the number of normal step sequence patterns relative to the total number of paw placements (Figure 1C) and the base of support of the hind paws (average width between the paws, Figure 1D) were not improved in injured mice treated with PRE-084. Similarly, σ1R agonist treatment had no effect on the print position of paws on the ipsilateral side of the spinal cord lesion (corresponding to the distance between the position of the hind paw and the position of the previously placed front paw in the same step cycle, Figure 1E). Also, the stride length and duty cycle on the ipsilateral side (indicating the stance duration as a percentage of the duration of the step cycle) remained unchanged with PRE-084 treatment (Figures 1F,G).
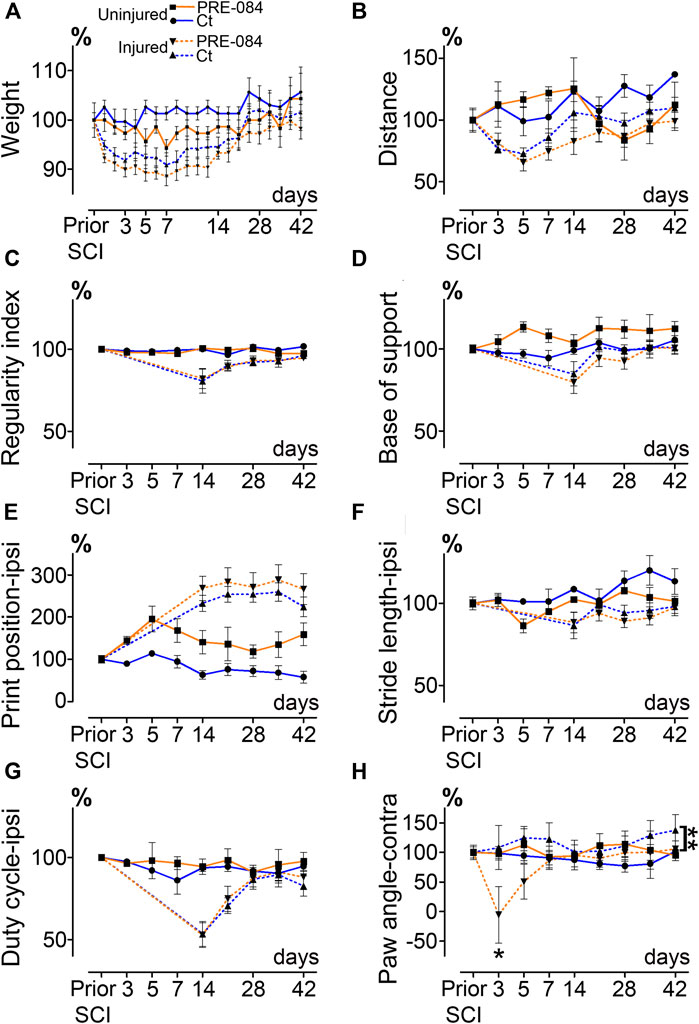
Effects of PRE-084 treatment on spontaneous motor activity and gait patterns after SCI.
Weight curves (A). Open field analysis of the distance covered in 8 min by mice in all groups (B). Walking pattern analysis using CatWalk XT®(C–H). Parameters analyzed include the regularity index (C) and the base of support (D). For paws located ipsilateral to the spinal cord lesion PRE-084 does not improve walking parameters such as the print position (E), the stride length (F) and the duty cycle (G). The paw angle of the hind paw located contralateral to the lesion is altered in the SCI-treated mice (H). Number of mice: Uninjured mice 6 (3 treated and three untreated, plain lines), hemisected mice 24 (12 treated and 12 untreated, dashed lines). Statistics: two-way ANOVA followed by Bonferroni tests in the comparison between injured control and injured treated groups (dashed lines), **p < 0.01.
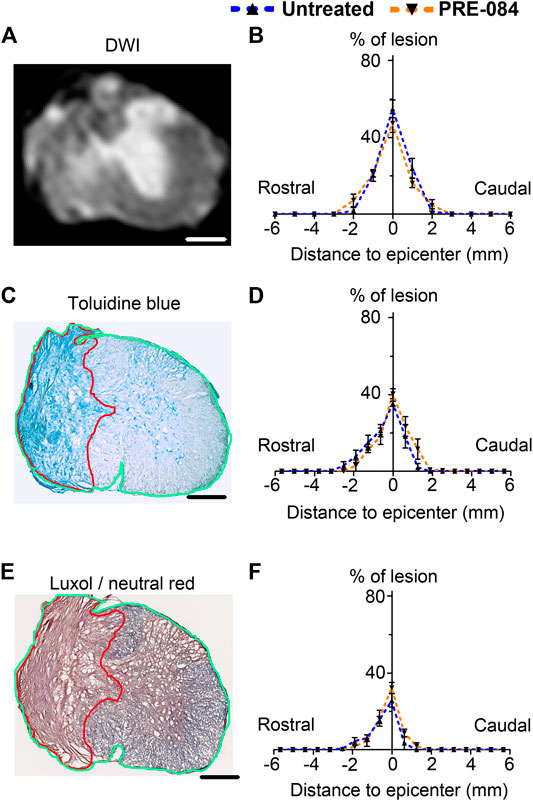
Lesion size analysis. Representative ex vivo diffusion weighted MRI (DWI)
(A), toluidine blue staining (C) and luxol fast blue/neutral red staining (E) at lesion epicenter 6 weeks after SCI. Quantifications of the lesion percentage at the epicenter, the lesion extension and the lesion volume represented by area under the curve using DWI acquisitions (B), toluidine blue sections staining (D) and luxol fast blue/neutral red (F) in both treated and untreated injured animals. Note the damaged spinal cord tissue (outlined in red in C and E) used for lesion quantification and allows distinction from undamaged tissue (outlined in green in C and E) Number of hemisected mice: DWI six treated and six untreated and histology five treated and five untreated. Scale bars: A, C AND E 600 μm.
On the contralateral side, σ1receptor activation transiently worsened contralateral hind paw placement since the paw angle (angle between the paw axis and the body axis) differed more from preoperative value (100% correspond to the angle prior to SCI) in the SCI-treated than in the SCI-untreated group (Figure 1H, p = 0.049). Muscle alterations can affect paw angle placement, we thus investigated the gastrocnemius-soleus-plantaris complex (Figure 3 and Supplementary Figure 1A). No difference in number of neuromuscular junction densities in between groups was observed (Figures 3A–C, ipsilateral p = 0.15; untreated: 5.07 ± 0.33; PRE-084: 4.14 ± 0.45 and contralateral p = 0.2; untreated: 4.91 ± 0.43; PRE-084: 4.13 ± 0.30), however when taking into account all sections that were quantified (and not only the mean per animal), PRE-084-treatment decreased the neuromuscular junction densities in both hind limbs (Supplementary Figure 1A, p < 0.0001 for both ipsilateral; untreated: 5.22 ± 0.11; PRE-084: 4.19 ± 0.12 and contralateral; untreated: 5.07 ± 0.11; PRE-084: 4.00 ± 0.09).
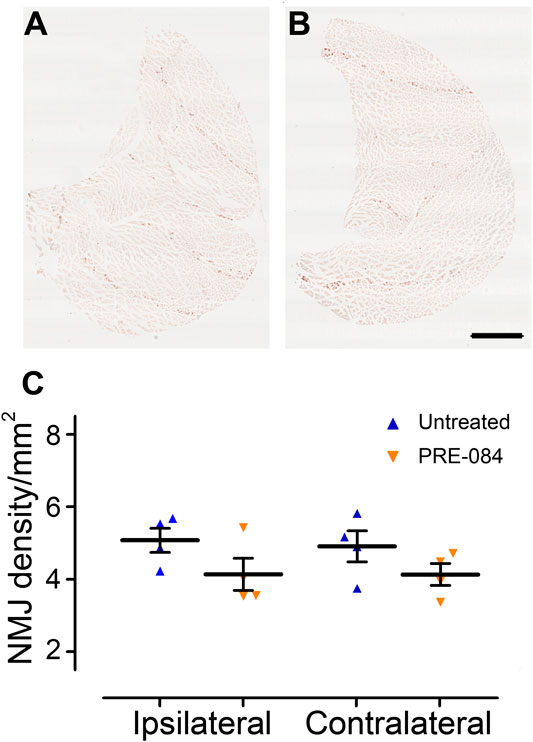
FIGURE 3.
Effects of PRE-084 treatment on the overall neuromuscular junction density.
Gastrocnemius–soleus–plantaris muscle complex of the hind limb located on the ipsilateral side of the lesion in untreated (A) and PRE-084-treated (B) mice. Quantitative assessments of neuromuscular junctions’ density in gastrocnemius–soleus–plantaris muscular complexes in untreated and PRE-084-treated mice (C). Each dot represents the mean of at least 15 sections counted per animal per muscle. Number of injured mice: four treated and four untreated. Scale bars A AND B 1 mm.
Therefore, σ1receptor activation over a 6-weeks period starting immediately after SCI is slightly detrimental to motor function recovery in mice.
σ1R Agonist Treatment After SCI Decreases NeuN Expression and Increases Astrocytic Reactivity Without Limiting Lesion Size
To analyze outcomes of σ1receptor activation on tissue structure following spinal cord lesion we acquired ex vivo DW-MRI (Figure 2A) and segmented 1 mm-thick axial slices. Lesion extension on the rostro-caudal axis, lesion volume and percentage of damaged tissues at lesion epicenter were similar in both groups (Figure 2B). We further assessed the effect of PRE-084 on spinal cord lesion size using classical histological methods such as toluidine blue (Figures 2C,D) and luxol fast blue/neutral red (Figures 2E,F) staining on the same spinal cord segment and confirmed the absence of difference between untreated and treated groups (Figures 2D,F). We then deepened our analysis and quantified the expression of neuronal (Figures 4A,B and a’&b’) and glial markers (Figures 4D,E and d’&e’; G&H and g’&h’; J&K and j’&k’) on a 1.2 cm-perilesional segment of the spinal cord using specific neuron (NeuN), astrocytes (GFAP), microglia and macrophages (IBA1) and microglia (TMEM 119) (Figures 4A–K) markers. We highlighted a decrease in NeuN expression in the treated group as compared to the untreated group in the dorsal horn of the rostral segment (p = 0.032; untreated: 12.37 ± 0.23; PRE-084: 11.00 ± 0.43) (Figure 4C), though when taking into account all sections that were quantified (and not only the mean per animal), PRE-084-treatment decreased NeuN expression in all regions (gray matter, ventral and dorsal horns in the rostral (p < 0.0001) and caudal (p < 0.05) segments (Supplementary Figure 1B). GFAP expression was higher in the white matter of the treated group caudal to the lesion (p = 0.048; untreated: 25.94 ± 0.82; PRE-084: 28.79 ± 0.85) (Figure 4F), similarly when taking into account all sections that were quantified, PRE-084-treatment increased GFAP expression in all regions rostral to the lesion (p < 0.0001) and in the gray and the white matters (with the exception of the dorsal funiculus) caudal to the injury (Supplementary Figure 1C, p < 0.0001). No modulation of microglia/macrophages was induced by σ1R agonist treatment. The overall IBA1 (Figure 4I and Supplementary Figure 1D) and TMEM119 (Figure 4L and Supplementary Figure 1E) expressions were similar in both groups.
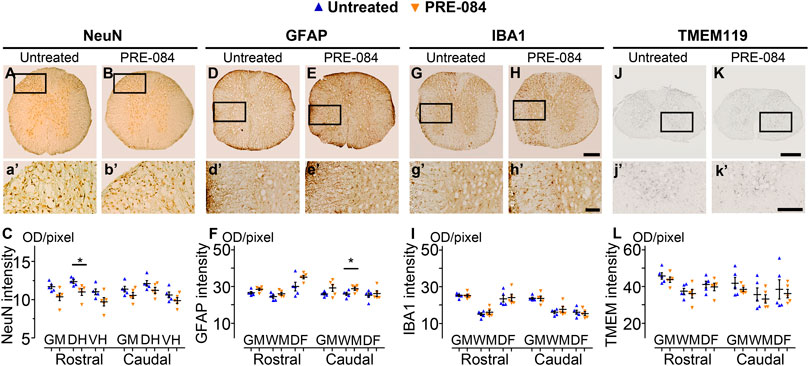
Histological analysis.
NeuN-positive neurons in the gray matter 6 weeks after SCI in untreated (A AND A’) and PRE-084-treated (B AND B’) mice rostral to the lesion site. Quantification of NeuN-immunoreactivity in the gray matter, the dorsal and the ventral horns, rostral and caudal to the lesion (C). GFAP-positive astrocytes 6 weeks after SCI in untreated (D AND D’) and PRE-084-treated (E AND E’) mice rostral to the lesion site. Quantification of GFAP-immunoreactivity in the gray matter, the white matter (excluding the dorsal funiculus) and the dorsal funiculus rostral and caudal to the lesion (F). IBA1-positive microglia/macrophage 6 weeks after SCI in untreated (G AND G’) and PRE-084-treated (H AND H’) animals rostral to the lesion site. Quantification of IBA1-immunoreactivity in the gray matter, the white matter (excluding the dorsal funiculus) and the dorsal funiculus, rostral and caudal to the lesion (I). TMEM 119-positive microglia 6 weeks after SCI in untreated (J AND J’) and PRE-084-treated (K AND K’) animals caudal to the lesion site. Quantification of TMEM119-immunoreactivity in the gray matter, the white matter (excluding the dorsal funiculus) and the dorsal funiculus rostral and caudal to the lesion (L). Higher magnifications (A’-B’, D’-E’, G’-H’ and J’-K’) of black insets in (A–B, D-E, G-H and J-K). In all graphs, results for untreated mice are in blue and PRE-084-treated are in orange. Data are expressed as mean ± SEM per group. Statistics: Student unpaired t-test with Welch correction (C, F, I AND L), *p < 0.05. Scale bars: A AND B, D AND E, G AND H 600 μm; J AND K500 μm; A’ AND B’, D’ AND E’, G’ AND H’ and J’ AND K’ 200 μm. GM: gray matter; VH: ventral horn; DH: dorsal horn; WM: white matter; DF: dorsal funiculus; OD = optical density. Each dot represents the mean of at least nine sections counted per animal per condition (630 μm interval between each section). Number of injured mice: five treated and five untreated.
Altogether, these results suggest that activation of σ1 receptor using PRE-084 over 6 weeks after injury induces a decrease in NeuN expression and an increase in astrogliosis while microglia reactivity is not affected.
Discussion
In this study, we assessed outcomes of sigma-1 receptor agonist treatment on functional recovery and tissue reorganization after SCI in mice. Our findings suggest a detrimental effect of PRE-084 post injury treatment on motor function recovery and is associated, at tissue level, with a decrease in NeuN expression and an increase in astrogliosis.
The involvement of σ1R in central neuropathic pain induced by SCI had been recently studied using both genetic and pharmacological approaches (Castany et al., 2018; Castany et al., 2019). Following spinal cord contusion at thoracic level 8–9 mice lacking σ1R (σ1KO) display reduced injury-induced thermal hyperalgesia and mechanical allodynia. Moreover, σ1KO do not present an injury-induced increase in spinal cord expression of the phosphorylated form of two molecules involved in central sensitization in neuropathic pain states i.e. extracellular signal-regulated kinases (ERK1/2) and NMDA receptor NR2B subunit. Conversely to wild type mice, SCI in σ1KO mice does not induce an upregulation of the pro inflammatory cytokines tumor necrosis factor (TNF-α) and interleukin IL-1β (Castany et al., 2018). Mice treated with a selective σ1R antagonist starting either 1 month (Castany et al., 2018) or 1 week (Castany et al., 2019) following SCI, similarly display reduced induced-central neuropathic pain. No major modification of motor activity was observed in both treatment protocols. Moreover, σ1R antagonist treatment during the first week post SCI also prevented injury-induced increases of pERK1/2, pNR2B-NMDA, TNF-α and IL-1β (Castany et al., 2019). In order to implement the two previous studies that investigated outcomes of a σ1R pharmacological modulation following SCI, we have chosen to perform our experiments in female mice. It had been shown a gender-dependent locomotor recovery following SCI; thus we cannot exclude a differential effect of sigma one agonist treatment in male mice (Farooque et al., 2006). Our findings, further suggest a detrimental effect of σ1R activation following SCI on motor function recovery that may thus result from an increased expression of proinflammatory molecules as reflected by an enhanced astrogliosis.
Sigma-1 receptor modulates calcium signaling between the endoplasmic reticulum and mitochondria and regulates mitochondrial functions, such as intramitochondrial Ca2+homeostasis, reactive oxygen species (ROS) generation and cellular bioenergetics (Hayashi and Su, 2007). Mitochondrial dysfunction is an essential component of the secondary injury cascade induced by SCI. In particular, it leads to ATP loss and successive inactivation of ATP-dependent ion pumps required for regulation of ion concentrations and reuptake of the excitatory neurotransmitter glutamate. Moreover, a consequence of the persistent ion shift during secondary injury is an increased ROS. On the one hand, sigma-1 receptor agonists favor brain plasticity and induce protective effect in the CNS under pathological conditions (Maurice and Goguadze, 2017) but on the other hand isolated mouse brain mitochondria exposed to σ1R agonists display an increase in ROS level. Strikingly, PRE-084 induces a ROS increase of 24% associated with a 40% selective increase of complex one activity when applied to isolated mouse brain mitochondria (non-pathological condition) and decreases Aβ1–42-induced ROS elevation (pathological condition i.e. amyloid toxicity) (Goguadze et al., 2019). Therefore, σ1R agonists exert condition-dependent effects on ROS production. The detrimental in vivo effect of per os PRE-084 treatment over 6 weeks at this dose (1 mg/kg/day) following lateral hemisection of the spinal cord may thus be a consequence of an increased ROS level.
The present study therefore suggests the detrimental effect of σ1R agonist on motor recovery, neuronal preservation and astrogliosis following SCI. However, dose-dependent and time response effects of σ1R agonists and antagonists treatments in various animal models of traumatic CNS injury merits further investigation