
Validation of σ2R/TMEM97 as a neuropathic pain target: Specificity, human expression and mechanism of action
By Muhammad Saad Yousuf, James J. Sahn, Eric T. David, Stephanie Shiers, Danielle M. Royer, Chelsea D. Garcia, Jennifer Zhang, Veronica M. Hong, Ayesha Ahmad, Benedict J. Kolber, Daniel J Liebl, Stephen F. Martin, and Theodore J. Price
Excerpt from the article published in bioRxiv 2023.04.11., 536439; DOI: https://doi.org/10.1101/2023.04.11.536439
Editor’s Highlights
- The sigma 2 receptor (σ2R) was recently identified in 2017 as transmembrane protein 97 (TMEM97).
- The analgesic effects of a σ2R/TMEM97 ligand in mice of both sexes require direct modulation of σ2R/TMEM97, not σ1R or any other protein or receptor.
- Modulation of σ2R/TMEM97 leads to the inhibition of integrated stress response (ISR) in mouse and human neurons.
- The ISR is a major mechanism that controls long-lasting transcriptional changes by influencing the translation of key injury-induced transcription factors like activating transcription factor 4 (ATF4) and C/EBP homologous protein (CHOP).
- σ2R/TMEM97 localization to the ER membrane may link it to the ISR via ER stress, particularly by influencing eIF2α phosphorylation by the kinase PERK.
- σ2R/TMEM97 is likely involved in transporting bioactive lipids, such as hydroxycholesterols, between cellular compartments, perhaps via a transporter activity like that of Niemann-Pick C1 (NPC1) protein.
- An “agonist-like” ligand of σ2R/TMEM97 promotes neurite outgrowth and complexity in a σ2R/TMEM97-dependent manner.
- Neuroregeneration is a protein-demanding process, and drugs that inhibit ISR, and thereby promote protein synthesis also enhance neurite outgrowth.
- Nerve injury and regeneration are inherently linked such that the mechanisms that govern neuropathic pain are inversely correlated with neuroregenerative processes.
Abstract
The Sigma 2 receptor (σ2R) was described pharmacologically more than three decades ago, but its molecular identity remained obscure until recently when it was identified as transmembrane protein 97 (TMEM97). We and others have shown that σ2R/TMEM97 ligands produce analgesia in mouse neuropathic pain models with a time course wherein analgesic onset is 24 hours following dosing. We sought to understand this unique anti-neuropathic pain effect by addressing two key questions: do these σ2R/TMEM97 compounds act selectively via the receptor, and what is their downstream mechanism on nociceptive neurons. Using male and female conventional knockout (KO) mice for Tmem97, we find that a novel σ2R/TMEM97 binding compound, FEM-1689, requires the presence of the gene to produce analgesia in the spared nerve injury model in mice. Using primary mouse dorsal root ganglion (DRG) neurons, we demonstrate that FEM-1689 inhibits the integrated stress response and promotes neurite outgrowth via a σ2R/TMEM97-specific action. We extend the clinical translational value of these findings by showing that FEM-1689 reduces ISR and p-eIF2α levels in human sensory neurons and that it alleviates the pathogenic engagement of ISR by methylglyoxal. We also demonstrate that σ2R/TMEM97 is expressed in human nociceptors and satellite glial cells. These results validate σ2R/TMEM97 as a promising target for further development for the treatment of neuropathic pain.
Significance Statement Neuropathic pain is a major medical problem that is poorly treated with existing therapeutics. Our findings demonstrate that σ2R/TMEM97 targeting with modulators creates analgesia in a mouse model via a specific action on the receptor. We also identify a potential mechanism of action, ISR inhibition, that links the receptor to cellular signaling events that have preclinical and clinical validation for pain relief. Our work suggests that σ2R/TMEM97 can be selectively engaged by specific small molecules to produce ISR inhibition in a subset of cells that are critical for neuropathic pain. σ2R/TMEM97-targeted therapeutics thus have the potential to offer effective pain relief without the side effects associated with currently available neuropathic pain medicines.
Introduction
Neuropathic pain, which is caused by an injury or disease of the somatosensory nervous system, affects approximately 10% of the population and is the leading cause of high-impact chronic pain (1). Management of neuropathic pain is a major clinical challenge because available drugs not only have limited efficacy, but they also elicit serious side effects. There is a significant need for novel drugs that alleviate neuropathic pain through non-opioid and non-addicting mechanisms and have improved side effect profiles.
The sigma 2 receptor (σ2R) was recently identified in 2017 as transmembrane protein 97 (TMEM97) (2). We discovered that several small molecules that bind selectively to σ2R/TMEM97 produce strong and long-lasting anti-neuropathic pain effects from spared nerve injury (SNI) in mice (3), a finding that was independently replicated with structurally distinct molecules (4). Although the biological function of σ2R/TMEM97 is not well understood, it is a transmembrane protein that is associated with the endoplasmic reticulum (ER) and plays a role in calcium signaling (5, 6) and cholesterol trafficking and homeostasis (7–11). 20(S)-Hydroxycholesterol was recently identified as an endogenous ligand for σ2R/TMEM97 (12). The role of σ2R/TMEM97 in disease pathology has been focused on cancer (13), but it is also implicated in neurodegenerative diseases including Alzheimer’s disease (14–17) and Parkinson’s disease (18). Pharmacological targeting of σ2R/TMEM97 has neuroprotective effects in a number of models of neurodegenerative conditions, including traumatic brain injury (19), Huntington’s disease (20), and retinal ganglion cell degeneration (21).
The mechanism by which modulation of σ2R/TMEM97 alleviates neuropathic pain is not known. Given the localization of σ2R/TMEM97 at the ER, the primary hypothesis tested in our work is whether σ2R/TMEM97 targeting may reduce pain via interference with the integrated stress response (ISR), which includes ER stress. The ISR is an adaptive response to cellular stressors such as accumulation of misfolded proteins, amino acid and heme deprivation, and viral infection (22, 23). A canonical signaling event associated with the ISR is phosphorylation of eukaryotic initiation factor 2α (eIF2α) in response to cellular stress conveyed by four kinases: protein kinase R (PKR), PKR-like ER kinase (10), heme-regulated inhibitor (HRI), general control nonderepressible 2 (GCN2). These kinases phosphorylate eIF2α, in turn, inhibiting global protein synthesis and promoting the translation of mRNAs with upstream open reading frames (uORF) like activated transcription factor 4 (ATF4). We and others have previously demonstrated that the induction of the ISR is associated with neuropathic pain caused by traumatic nerve injury (24, 25), metabolic disorders (26–28), and autoimmune disorders (29–31).
Another key question is whether the analgesic effects of σ2R/TMEM97 ligands is specifically due to their binding to σ2R/TMEM97 because such compounds can also have substantial activity at the sigma 1 receptor (σ1R), a receptor that also produces analgesia in animal models (32–35). We used a recently developed knockout mouse of the Tmem97 gene, and a new small molecule, FEM-1689, that has improved selectivity for σ2R/TMEM97 to test the hypothesis that σ2R/TMEM97 is causatively linked to analgesia in mouse neuropathic pain models. Indeed, the analgesic effect of FEM-1689 in mouse neuropathic pain is completely absent in TMEM97KO mice. Our findings also show that FEM-1689 inhibits the ISR in a σ2R/TMEM97-dependent fashion in mouse and human DRG neurons. This work provides a strong mechanistic case for targeting of σ2R/TMEM97 as the basis of a new approach to treat neuropathic pain.
Results
TMEM97 mRNA is expressed in the human and mouse dorsal root ganglia
To assess whether σ2R/TMEM97 is expressed in human nociceptors, we performed RNAscope in situ hybridization using human dorsal root ganglia (DRG) obtained from organ donors. We found that TMEM97 is expressed in all classes of human sensory neurons including SCN10A (Nav1.8)-positive, putative nociceptors (Fig 1A-C). Approximately 70% of all neurons expressed SCN10A transcripts, and all neurons evaluated expressed TMEM97(Fig 1C). The average diameter of neurons expressing TMEM97 was roughly 74 µm as compared to SCN10A-expressing nociceptors that are 48 µm (Fig 1D). Upon further investigation, we found that FABP7-positive satellite glial cells also expressed TMEM97 (Fig 1E). We validated our findings that TMEM97 is expressed across all neuronal subtypes in the human DRG concurrent with our previously published human DRG spatial RNA sequencing dataset (36). Mouse DRG neurons and satellite glial cells also express Tmem97 mRNA (Supp Fig 1A) with a nearly identical pattern to what was seen in the human DRG. Moreover, previously published single-cell RNA sequencing data, from mice shows that Tmem97 is expressed in all DRG neuronal subtypes including Scn10a-positive nociceptors, and Nefh-positive large diameter proprioceptors and low-threshold mechanoreceptors (Supp Fig 1B) (37).
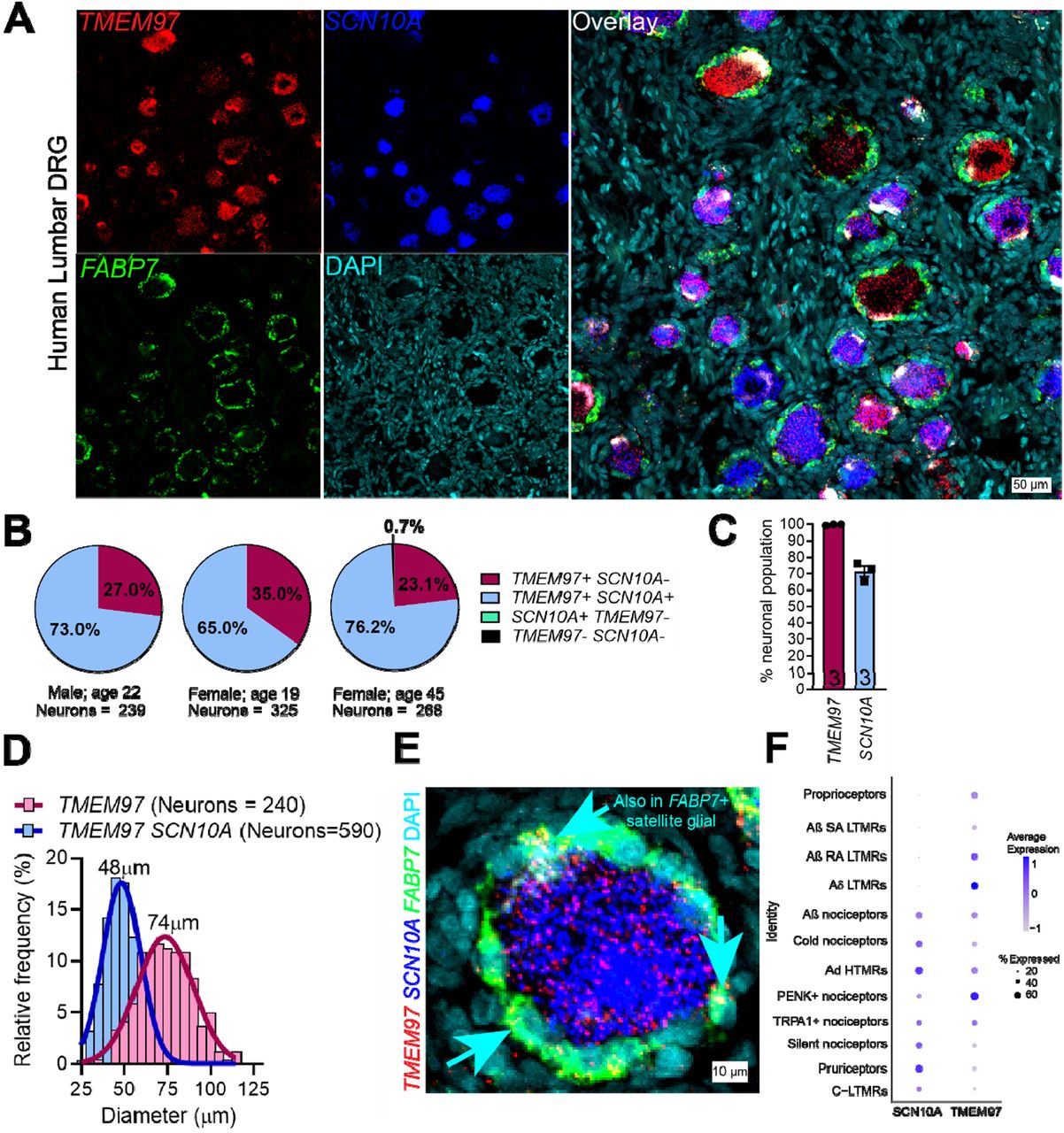
TMEM97 gene is expressed in human dorsal root ganglia (DRG).
(A) RNAScope in situ hybridization experiments using lumbar DRGs obtained from organ donors. (B-C) Across 3 donors (1 male and 2 females), we discovered that nearly all DRG neurons (>99%) expressed TMEM97 and notably, all SCN10A-positive nociceptors expressed TMEM97. (D) TMEM97-positive neurons were distributed across all cell sizes. (E) Upon further investigation, we also identified TMEM97 transcripts in FABP7-positive satellite glial cells. (F) Our previously published (36) analysis of near single-cell RNA sequencing of human DRGs showed that TMEM97 is expressed across all neuronal cell types in the ganglia including nociceptors, low-threshold mechanoreceptors (LTMRs), and proprioceptors.
Discovery of FEM-1689 as a potent σ2R/TMEM97 binding ligand
Methanobenzazocines and norbenzomorphans represent two distinct chemotypes of biologically active ligands that bind selectively to σ2R/TMEM97 (Fig 2) (38, 39). The first group comprises UKH-1114, which alleviates mechanical hypersensitivity in a mouse model of neuropathic pain (3) and reduces neuronal toxicity in a model of Huntington’s disease (20). Based on the positive outcomes using UKH-1114 as a treatment in models of neuropathic pain, we modified its chemical structure and synthesized FEM-1689 (Supp Fig 2), which is a highly selective compound with improved pharmacokinetic and physicochemical properties (Fig 3). FEM-1689 is >100-fold more selective for σ2R/TMEM97 than 40 CNS proteins except for σ1R (19-fold) and norepinephrine transporter (NET; 37-fold) (Supp Table 1).
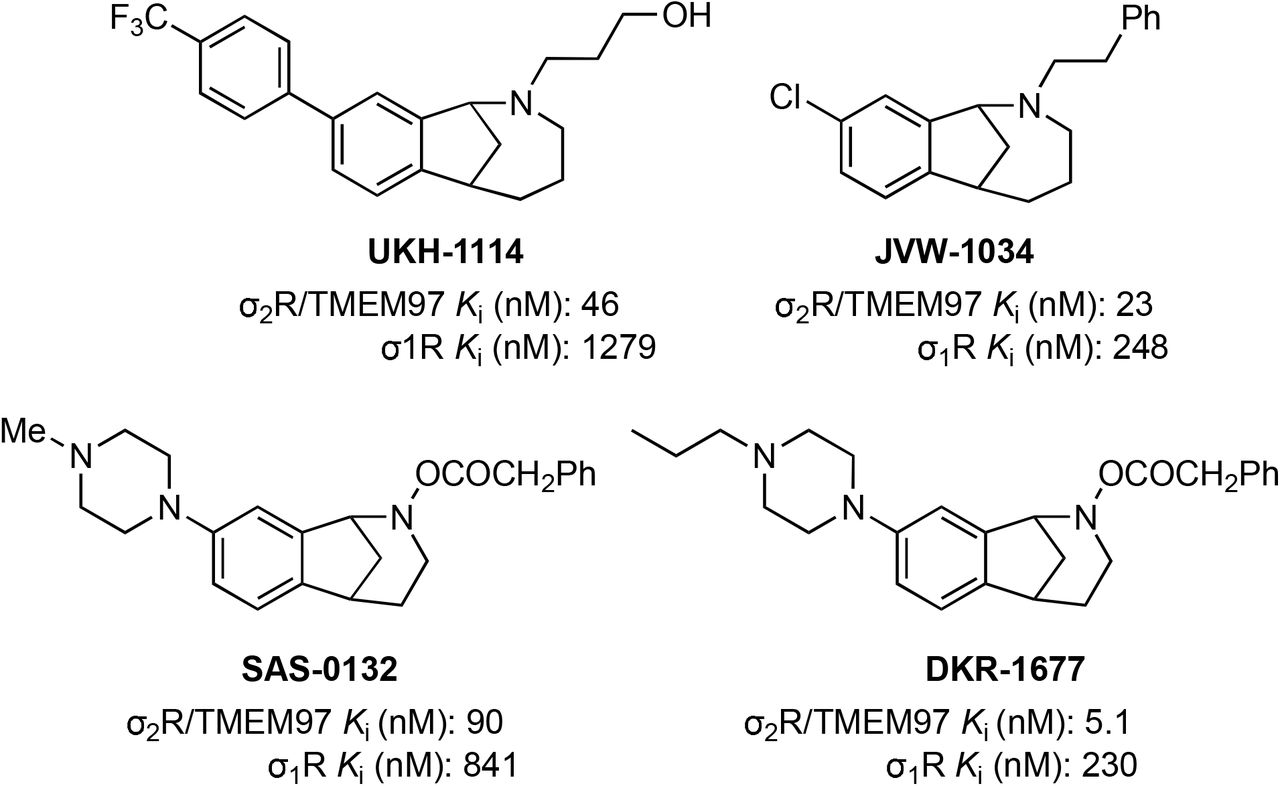
Structures and binding affinities of selected biologically-active methanobenzazocines (e.g. UKH-1114 and JVW-1034) and norbenzomorphans (e.g. SAS-0132 and DKR-1677). Ki values were determined at the PDSP using σ2R/TMEM97 sourced from rat PC12 cells and σ1R sourced from guinea pig brain, and values are reported as an average of two or more independent experiments (68).
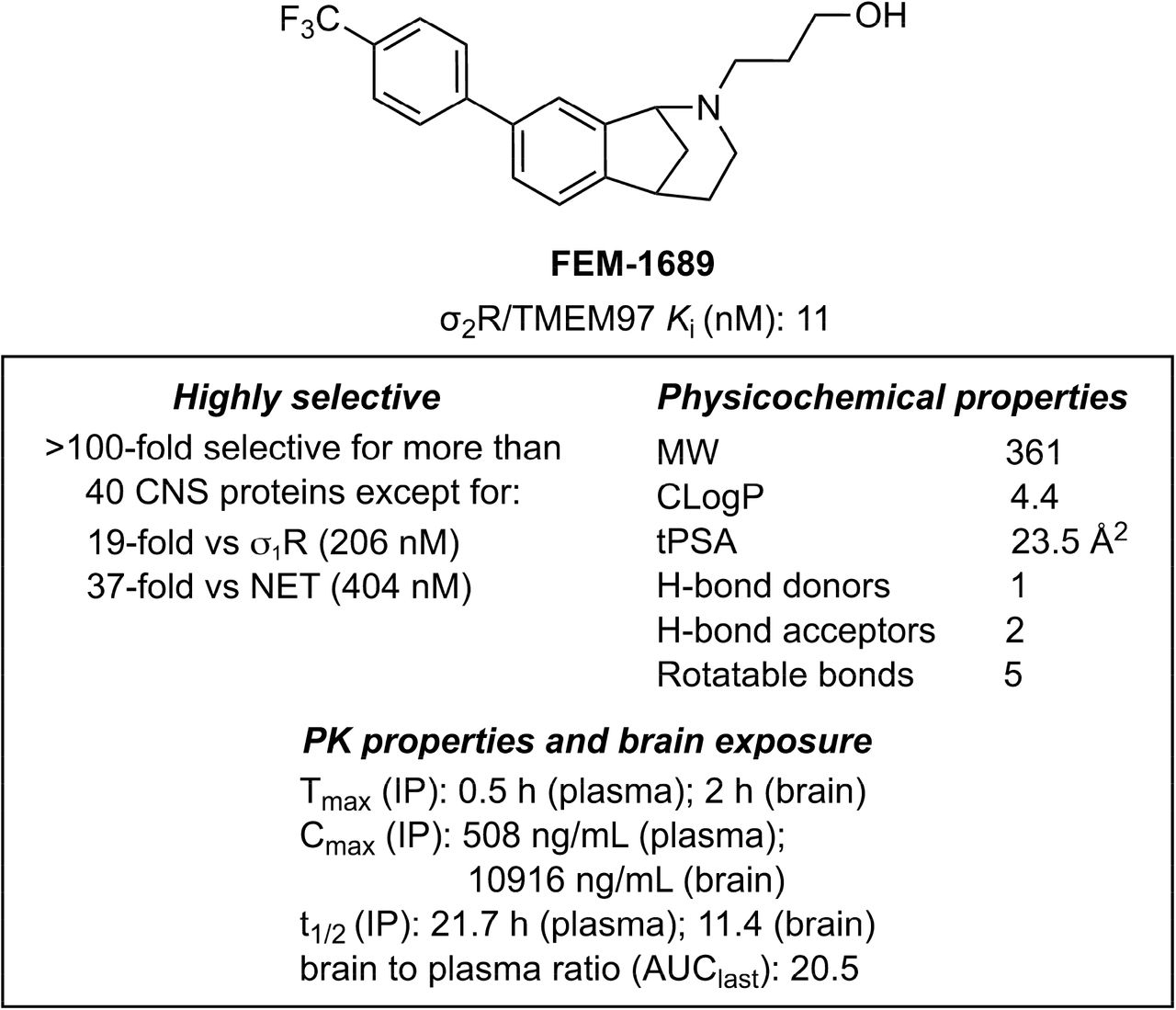
Structure, binding profiles, physicochemical properties, and pharmacokinetic parameters for FEM-1689.
Analgesic effect of FEM-1689 in male and female mice requires an intact Tmem97 gene
An important unresolved issue is whether σ2R/TMEM97 ligands promote analgesia specifically through action on σ2R/TMEM97. To address this question, we used a global TMEM97-knockout (KO) mouse. Male and female TMEM97KO animals and their wild-type counterparts had similar von Frey, Hargreaves’, and acetone responses suggesting that the loss of TMEM97 did not alter their baseline evoked mechanical, heat, and cold sensitivity (Supp Fig 3). We then examined whether TMEM97KO animals develop an enhanced or blunted mechanical neuropathic pain phenotype following spared nerve injury (SNI) (40) (Fig 4A). Male and female TMEM97KO and wild-type animals developed severe and prolonged mechanical hypersensitivity following SNI tested at days 7, 10, and 14 after nerve injury (Fig 4B, C). Animals were tested thirty days following nerve injury and were found to be still mechanically hypersensitive. Male and female wild-type and TMEM97KO animals were then treated with a single intravenous injection of FEM-1689 dosed at 10 mg/kg, a dose determined based on previous studies with another σ2R/TMEM97 ligand, UKH-1114 (3). We then assessed TMEM97KO and wild-type evoked mechanical threshold daily for 7 days and found little to no change in either group for mechanical sensitivity after 10 mg/kg of FEM-1689 (Fig 4D, E). Animals were allowed to recover for two weeks before a second treatment with a higher 20 mg/kg dose of FEM-1689. We found that a higher dose of 20 mg/kg was effective at reversing mechanical hypersensitivity in both males and female wild-type mice for roughly 4 days. Notably, FEM-1689 failed to produce analgesia in TMEM97KO mice suggesting that FEM-1689’s effects were dependent on TMEM97. The effect size measurements for FEM-1689 were quantified for males (Fig 4F) and females (Fig 4G).
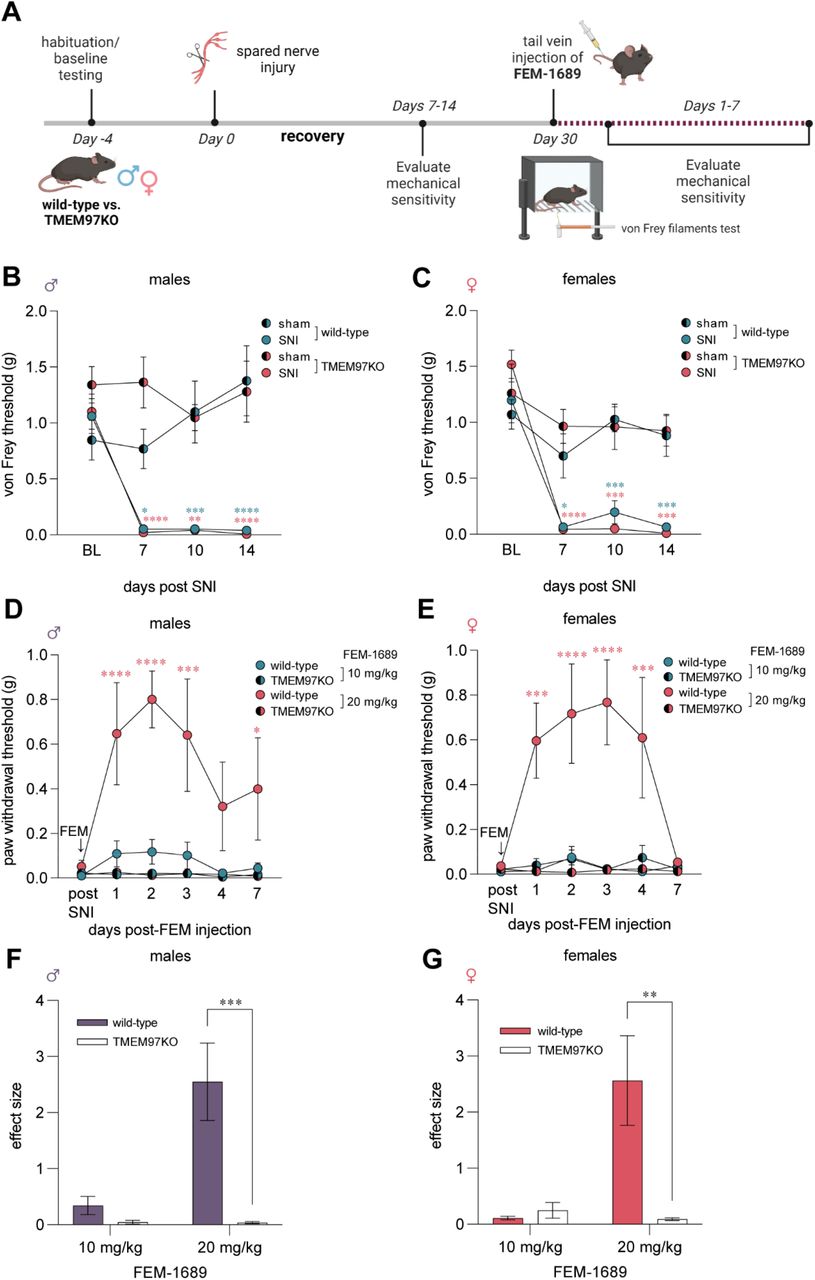
FEM-1689 inhibits the ISR and promotes neurite outgrowth in a σ2R/TMEM97-dependent fashion
We then sought to identify the mechanism by which FEM-1689 acts on sensory neurons through σ2R/TMEM97. Prior work has identified a link between σ2R/TMEM97 and cholesterol synthesis and trafficking, processes that are heavily regulated by 5’ adenosine monophosphate-activated protein kinase (AMPK) and its substrate, acetyl-CoA carboxylase (41). AMPK agonists are also known to produce analgesia in mouse models (22, 42–45). We treated cultured mouse DRG neurons with FEM-1689 at concentrations covering a range (10, 100, and 1000 nM) consistent with target binding (Ki = 11 nM) over the course of 16 hours and found no change in p-ACC levels whereas A769662 (100 µM), a known AMPK activator, increased p-ACC levels in both wild-type and TMEM97KO neurons (Supp Fig 4 A, B).
We then tested the hypothesis that FEM-1689 treatment might inhibit the ISR. Multiple lines of evidence support this hypothesis: 1) σ2R/TMEM97 and ISR transducers like protein kinase R-like ER kinase (PERK) are located on the ER membrane (39, 46); 2) a recent report on the effect of 20(S)-hydroxycholesterol on σ2R/TMEM97 implicated the ER-golgi network (12); and 3) the ISR is engaged in trauma-induced and diabetic neuropathic pain conditions (25, 26). We cultured mouse DRG neurons from wild-type and TMEM97KO animals, treated them with FEM-1689 over 16 hours, and measured changes in the levels of p-eIF2α using immunohistochemistry (Fig 5A, B). Interestingly, basal levels of p-eIF2α were much lower in TMEM97KO neurons than their wild-type counterparts. P-eIF2α levels following ISRIB treatment in wild-type neurons were comparable to p-eIF2α immunoreactivity in vehicle and ISRIB treated TMEM97KO neurons (Fig 5C). FEM-1689 reduced p-eIF2α levels, as compared to vehicle-treated cells, in wild-type mouse DRG neurons but not in DRG neurons cultured from TMEM97KO animals (Fig 5D, E). We found that FEM-1689 at 100 nM inhibited the ISR to the same extent as ISRIB (200 nM), a well-known ISR inhibitor (Fig 5D) (47). We calculated the p-eIF2α IC50 of FEM-1689 to be 30 nM in wild-type mouse DRG neurons, which was very similar to the binding affinity of FEM-1689 to TMEM97 (Ki=11 nM). FEM-1689 did not reduce levels of BiP, a chaperone important for initiating ER stress and ISR, in either wild-type or TMEM97KO neurons (Supp Fig 4C, D). Sholl analysis of cultured DRG neurons showed that FEM-1689 promoted neurite outgrowth in wild-type neurons but not in TMEM97KO cells (Fig 5F-K). Notably, neurite outgrowth in vehicle-treated TMEM97KO neurons was more pronounced than vehicle-treated wild-type neurons – an observation that may be due to reduced ISR in TMEM97KO cells and hence, enhanced protein synthesis. These data suggest that σ2R/TMEM97 is necessary for FEM-1689 to reduce ISR and promote neurite outgrowth.
FEM-1689 reduced p-eIF2α levels and promoted neurite outgrowth in vitro. (A, B) Cultured mouse DRG neurons obtained from wild-type and TMEM97KO animals were treated with FEM-1689 over 16 hours. Basal levels of p-eIF2α were lower in TMEM97KO neurons. ISRIB (200 nM) reduced p-eIF2α levels in wild-type neurons but failed to change p-eIF2α immunoreactivity in TMEM97KO neurons (C). Immunoreactivity of p-eIF2α was assessed and a 9-point dose-response curve ranging from 0.1 nM to 1000 nM was generated as a percentage of p-eIF2α levels of vehicle treated cells. An IC50 of 30 nM was determined with maximal effect at 100 nM which was comparable to ISRIB (200 nM) treatment (D, E). TMEM97KO DRG neurons did not respond to FEM-1689 treatment. (F-K) Sholl analysis of mouse DRG neurons following 100 nM of FEM-1689 treatment showed an increase in the number and complexity of neurites in wild-type neurons but not in TMEM97KO neurons. Area under the curve of Sholl analysis was used to statistically demonstrate this effect. Immunoreactivity against β3-tubulin was used to identify neuronal cell bodies and neurites. ***p<0.001, ****p<0.0001 two-tailed Student’s t-test. UND=undetermined. &=average mean gray intensity of the primary omission control.
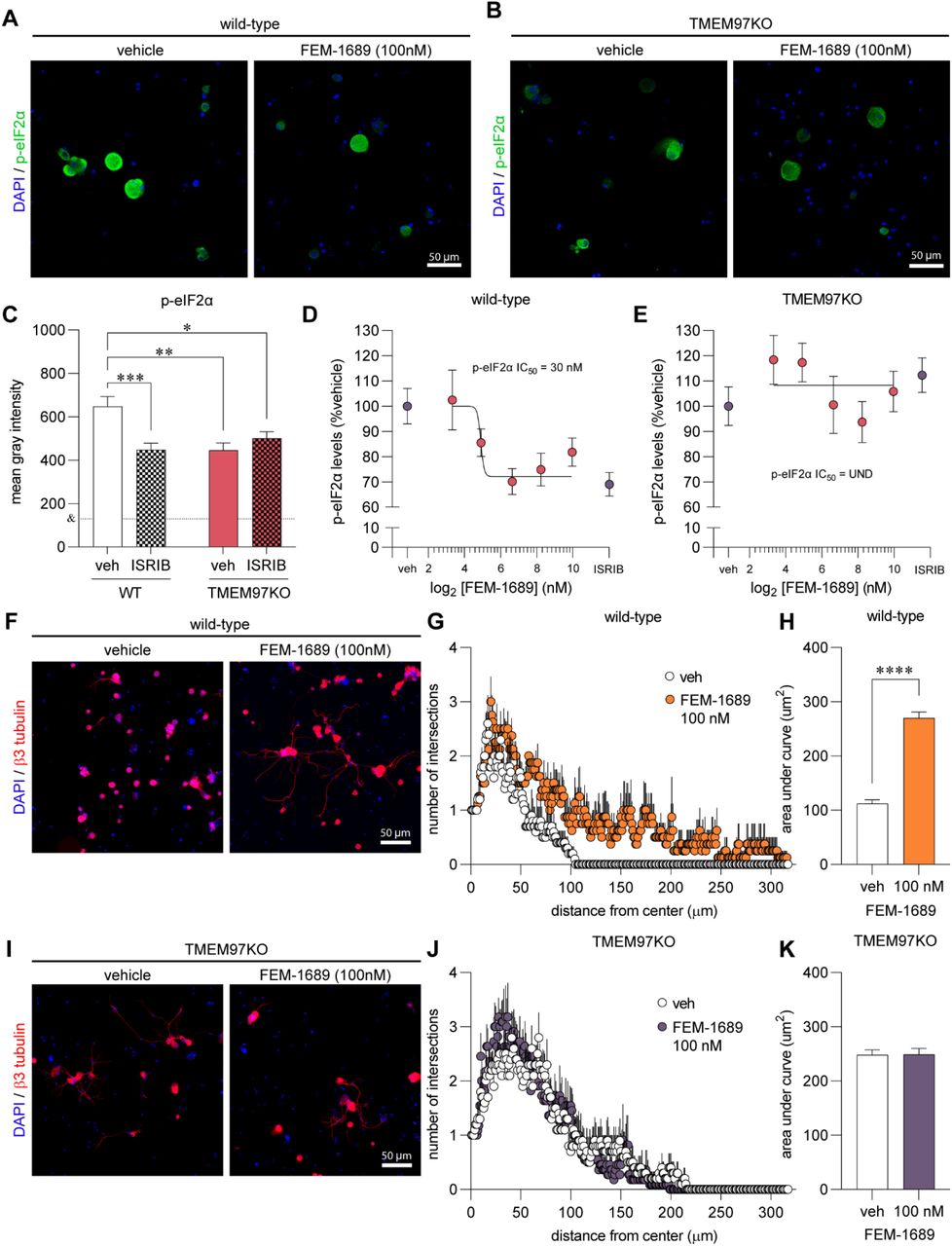
Norbenzomorphan σ2R/TMEM97 ligands SAS-0132 and DKR-1677 enhance the ISR
We then assessed whether other compounds that bind to σ2R/TMEM97 inhibit the ISR and whether ISR inhibition is specific to σ2R/TMEM97 modulators that promote analgesia. The norbenzomorphans FEM-1689, SAS-0132 and DKR-1677 and methanobenzacocines UKH-1114 represent two structurally distinct chemotypes that interact with the binding site of σ2R/TMEM97 in different orientations (39). SAS-0132 was previously shown to inhibit the analgesic effects of UKH-1114, which is a homolog of FEM-1689, suggesting that these two classes of molecules may exert opposing effects on σ2R/TMEM97 biology. After treatment of wild-type mouse DRG neurons with 10, 30, 100, and 300 nM of SAS-0132 and DKR-1677 over 16 hours, we found that both compounds promoted the phosphorylation of eIF2α, in contrast to the inhibitory effects of FEM-1689 (Fig 6).
Norbenzomorphans such as SAS-0132 (A-C) and DKR-1677 (D-F) stimulate the ISR by promoting the phosphorylation of eIF2α in cultured wild-type mouse DRG neurons. Cells were treated for 16 hours with either SAS-0132 or DKR-1677 at 10, 30, 100, or 300nM concentrations. Data is presented as fold-change compared to the fluorescence measured in the vehicle-treated neurons. *p<0.05, **p<0.01, ****p<0.0001 One-way ANOVA with Tukey’s post hoc test.
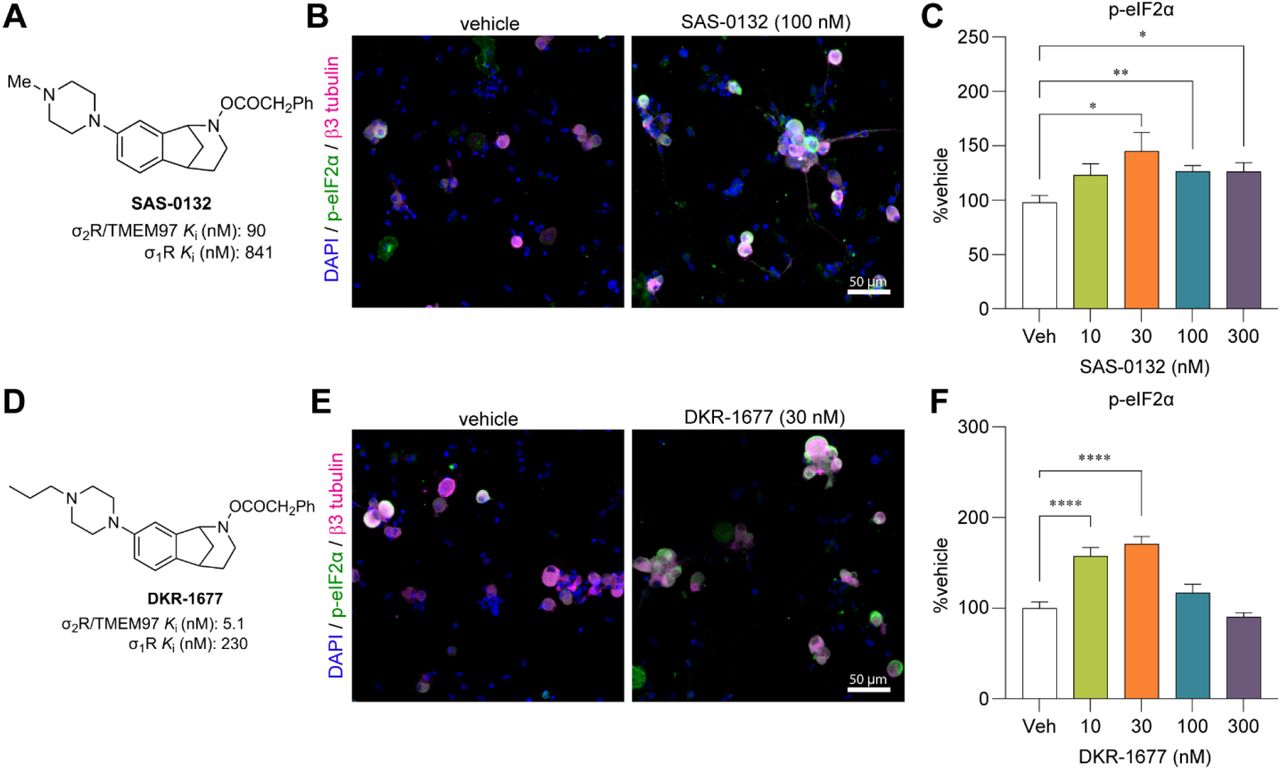
FEM-1689 inhibits the ISR in HEK293T cells
Human embryonic kidney (HEK) 293T cells express σ2R/TMEM97 (The Human Protein Atlas (48)), so we queried whether FEM-1689 would inhibit ISR in this cell line. We demonstrate a concentration-dependent reduction in p-eIF2α immunoreactivity following FEM-1689 treatment using immunocytochemistry (ICC) and spectrophotometry (Supp Fig 5A, B). We tested the effect of FEM-1689 over nine concentrations, ranging from 0.1 nM to 1000 nM and measured the p-eIF2α IC50 of FEM-1689 in HEK cells to be 0.9 nM (Supp Fig 5B). We further examined a broader panel of ISR-related proteins with increasing concentrations of FEM-1689 using Western blotting. HEK293T cells treated with FEM-1689 over 16 hours showed a concentration-dependent reduction in phosphorylation of eIF2α, eIF2A, and p-PERK, while BiP expression remained relatively stable throughout (Supp Fig 5C-K).
FEM-1689 reduces p-eIF2α and reverses MGO-induced ISR activity in human sensory neurons
To extend and translate our rodent findings to humans, we treated cultured human sensory neurons from organ donors with FEM-1689 for 16 hours. Consistent with our mouse data, we discovered that FEM-1689 significantly reduced p-eIF2α levels in human neurons at concentrations of 10 and 100 nM (Fig 7A, B). We then assessed the potential of FEM-1689 to reverse pathological ISR activation in human cells. To induce ISR in vitro, we treated human sensory neurons with methylglyoxal (MGO, 1 µM) – a metabolic by-product of glycolysis that is implicated in diabetic neuropathic pain and other painful neurodegenerative disorders. We have previously demonstrated that MGO induces an ISR response that causes mechanical hypersensitivity in rodents (26). MGO treatment increased p-eIF2α levels in human DRG neurons and co-treatment with FEM-1689 (100 nM) prevented the increase in p-eIF2α levels (Fig 7C, D). These findings support the conclusion that ISR activation associated with stimuli that cause neuropathic pain in humans can be blocked by TMEM97 modulation.
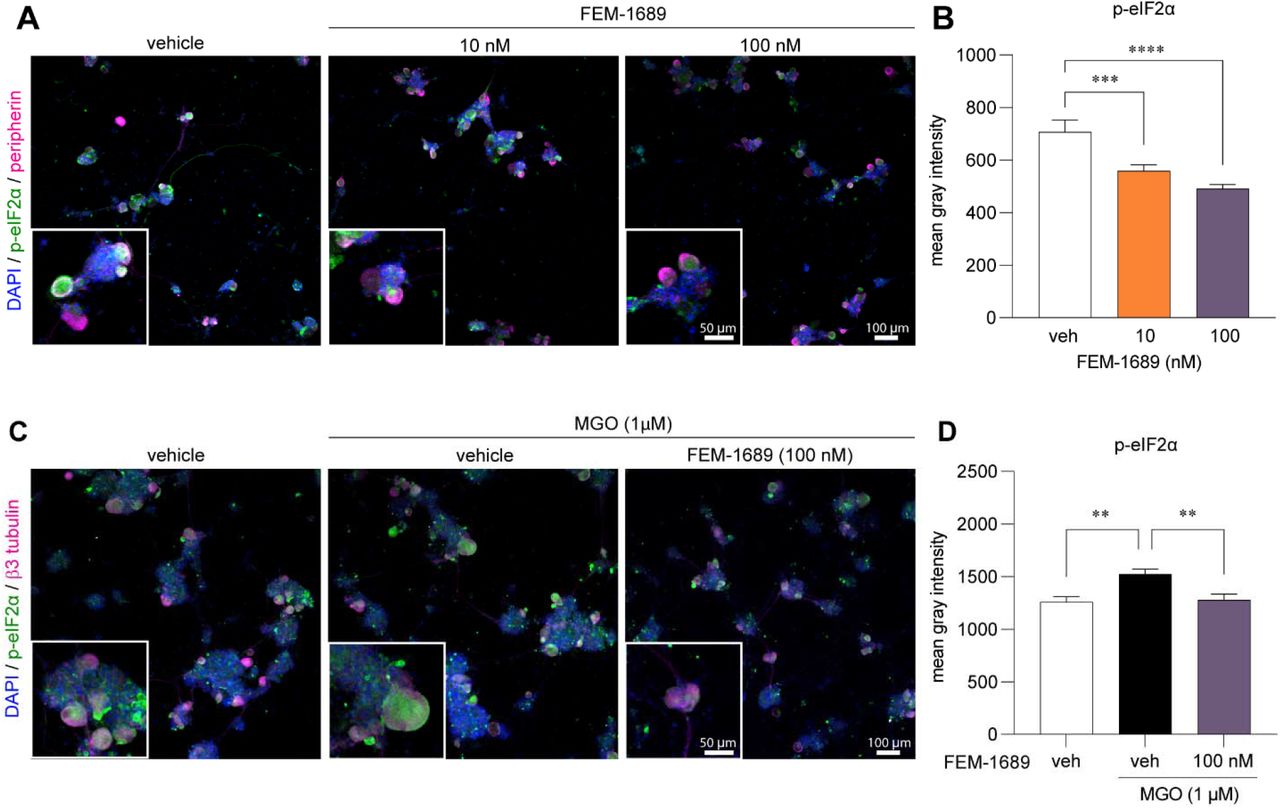
(A, B) FEM-1689 treatment (10 and 100nM) of cultured human DRG neurons significantly reduced p-eIF2α levels. (C, D) Methylglyoxal (MGO) is known to induce the ISR. Co-treatment of human neurons with MGO (1 µM) and FEM-1689 (100 nM) prevented an increase in p-eIF2α suggesting that FEM-1689 limits the effect of MGO. Peripherin and β3-tubulin were used to identify neurons. One-way ANOVA followed by Tukey’s post-hoc test **p<0.01, ***p<0.001, ****p<0.0001.
Discussion
Our experiments clearly demonstrate that the analgesic effects of a σ2R/TMEM97 ligand in mice of both sexes require direct modulation of σ2R/TMEM97, not σ1R or any other protein or receptor. This work also shows that modulation of σ2R/TMEM97 leads to ISR inhibition in mouse and human neurons. Insofar as a reduction in ISR has previously been linked to pain relief (22, 26, 29), we believe it is a plausible cellular mechanism for the analgesia that is caused by FEM-1689. Finally, we show that human nociceptors express the TMEM97 gene, FEM-1689 reduces eIF2α phosphorylation in cultured human sensory neurons and that the MGO-induced ISR in human sensory neurons can be reversed using FEM-1689. These observations suggest that targeting TMEM97 in pain patients could produce analgesia by inhibiting the ISR, a mechanism similarly observed in mice. We conclude that these findings validate σ2R/TMEM97 as a bona fide target for neuropathic pain development.
The time course of action of σ2R/TMEM97 modulators in mouse neuropathic pain models is different from many other analgesic compounds that have a rapid onset of action in accordance with the pharmacokinetics of the compound. Prior to this work, the slow onset time course of these analgesic effects in the mouse SNI model were independently observed by two different groups using distinct classes of σ2R/TMEM97 ligands (3, 4). Here, we have shown that this effect is mediated specifically by σ2R/TMEM97 because the antinociceptive activity is completely lost in male and female Tmem97 knockout mice. These observations suggest that the kinetics of signaling for σ2R/TMEM97 may be the primary underlying reason for the delayed analgesic response. The ISR is a major mechanism that controls long-lasting transcriptional changes by influencing the translation of key injury-induced transcription factors like activating transcription factor 4 (ATF4) and C/EBP homologous protein (CHOP). ISR mediated changes involving multiple transcriptional and post-translational modifications take time to initiate and progress. It is possible that this time course is responsible for the delay in the temporal action of FEM-1689 versus the pharmacodynamics of ligand binding to σ2R/TMEM97. The molecular signaling pathway downstream of σ2R/TMEM97 activation requires further investigation.
Over the course of our studies to discover compounds that bind selectively to σ2R/TMEM97, we have discovered several chemotypes that exhibit beneficial effects in a number of animal models (39). One group comprises aryl-substituted methanobenzazocines such as UKH-1114 and JVW-1034 (Fig 2). UKH-1114 alleviates mechanical hypersensitivity following nerve injury (3), and it reduces neuronal toxicity induced by mutant huntingtin protein in a model of Huntington’s disease (20). The methanobenzazocine JVW-1034 not only reduces withdrawal behaviors in two rodent models of alcohol-dependence (49, 50), but it also alleviates heightened pain sensitivity that is induced by chronic alcohol exposure in mice (50). Herein we report that FEM-1689, a close analog of UKH-1114, also alleviates mechanical hypersensitivity following nerve injury. Another structural class of σ2R/TMEM97 modulators include piperazine-substituted norbenzomorphans such as SAS-0132 and DKR-1677 (Fig 2). For example, SAS-0132 is neuroprotective and improves cognitive performance in animal models of age-related neurodegeneration (17, 51). Notably, SAS-0132 also blocks the antinociceptive activity of UKH-1114 (3), suggesting different σ2R/TMEM97 binding compounds may have opposing effects on analgesia. DKR-1677, a homolog of SAS-0132, is protective in two different models of traumatic brain injury (TBI). It reduces axonal degeneration and provides dose-dependent enhancement of cognitive performance in the blast injury model of TBI, while it protects oligodendrocytes and cortical neurons in the controlled cortical impact model (19). DKR-1677 also protects retinal ganglion cells from ischemia/reperfusion injury (21). We demonstrate herein that SAS-0132 and DKR-1677 increase p-eIF2α expression in mouse DRG neurons and have the opposite effect on ISR as compared to FEM-1689. This observation is further substantiated by the observation that A011, an unrelated σ2R/TMEM97 ligand, enhances ISR and promotes apoptosis in MCF-7 cells (52). These data show that σ2R/TMEM97 modulators can have opposing effects on ISR and that a reduction in ISR via modulation of σ2R/TMEM97 provides a screening framework for drug discovery for novel analgesics targeting this receptor.
Understanding how σ2R/TMEM97 modulators affect the ISR requires further investigation, but there are various clues for direct and indirect influence on the ISR in the literature. Firstly, σ2R/TMEM97 localization to the ER membrane may link it to the ISR via ER stress, particularly by influencing eIF2α phosphorylation by the kinase PERK. Indeed, we observed a reduction in p-PERK in HEK cells following FEM-1689 treatment at concentrations where p-eIF2α and eIF2A are maximally reduced (Supp Fig 5), suggesting a possible link between σ2R/TMEM97 and PERK. Second, σ2R/TMEM97 is likely involved in transporting bioactive lipids, such as hydroxycholesterols (12), between cellular compartments, perhaps via a transporter activity like that of Niemann-Pick C1 (NPC1) protein (9). Excessive lipid intake and the demand for increase lipid synthesis promote lipid stress of the ER that is known to activate the PERK-eIF2α branch of the ISR and impair mitochondrial function (53–55). Finally, σ2R/TMEM97 regulates cellular Ca2+ dynamics via its influence on store-operated calcium entry (11). The ER is the largest Ca2+ store in the cell and is sensitive to fluctuations in Ca2+ levels causing ER stress and the ISR (56). It is currently unclear whether one or multiple mechanisms are required to produce analgesia associated with FEM-1689 and further investigation is required.
Neurite outgrowth can be used to assess the neuromodulatory, neuroprotective, and neuroregenerative effects of drugs (57). Our data demonstrate that FEM-1689 promotes neurite outgrowth and complexity in a σ2R/TMEM97-dependent manner as measured by Sholl analysis. Neurons lacking σ2R/TMEM97 have enhanced neurite outgrowth compared to their wild-type counterparts under naïve conditions. The reason for this may be due to a reduced ISR (Fig 5C), and hence uninhibited protein synthesis in TMEM97KO neurons at basal levels. Neuroregeneration is a protein-demanding process, and drugs that inhibit ISR, like ISRIB, and thereby promote protein synthesis also enhance neurite outgrowth (58). TMEM97KO neurons have reduced p-eIF2α levels at baseline, comparable to levels of p-eIF2α in ISRIB-treated wild-type neurons. ISRIB treatment of TMEM97KO neurons does not further reduce p-eIF2α levels. This may represent a physiological floor effect where the ISR is inhibited maximally in TMEM97KO cell without being cytotoxic.
Nerve injury and regeneration are inherently linked such that the mechanisms that govern neuropathic pain are inversely correlated with neuroregenerative processes. Various neuropathic pain models, like SNI, diabetic neuropathy, and chemotherapy-induced neuropathy are characterized by reduced intraepidermal nerve fiber density while reinnervation into the epidermis in these models can alleviate pain hypersensitivity (59). Of note however aberrant innervation of the skin can also promote pain hypersensitivity suggesting that neuroregeneration is a balancing act between protective and detrimental mechanisms. We propose that modulating σ2R/TMEM97 promotes appropriate neurite regrowth and enhances protective, analgesic regenerative processes.
There are limitations to our work. First, we have not shown a definitive link between ISR activation and analgesia in vivo with FEM-1689. One possibility would be to evaluate neuropathic pain in Eif2s1-knockout mice. Such an experiment is complicated because loss of eIF2α, encoded by Eif2s1, or completely preventing the phosphorylation of eIF2α through a point mutation is lethal (60). Moreover, no specific antagonists of eIF2α have been described. The widely used classes of compounds that inhibit ISR are chemical chaperones, ISR kinase inhibitors, and eIF2B-stabilizers like ISRIB do not directly target eIF2α and are not appropriate tools to answer this question. Second, despite the strong connection we have shown between FEM-1689 and σ2R/TMEM97 for SNI-induced mechanical hypersensitivity, we fail to see any change in the nerve injury phenotype in TMEM97KO mice. This may point to an “agonist-like” action of FEM-1689. That is, the TMEM97KO (a permanent “antagonist”) mice would be expected to show an increase in pain but all of the animals are at a floor after SNI and their mechanical thresholds are technically indistinguishable. Third, we have not determined whether the activity of FEM-1689 is due to a peripheral or central site of action. While our pharmacokinetic experiments show that the compound readily enters the brain (Fig 2), we have shown a clear action on the DRG. Future experiments will use a cell-specific TMEM97 knockout mouse under development to solve this important issue.
We have validated σ2R/TMEM97 as a neuropathic pain target and identified FEM-1689 as a candidate molecule for optimization for further development. The recent report of the crystal structure of σ2R/TMEM97 bound with several ligands will facilitate advancements in σ2R/TMEM97 structure-affinity relationships that can be leveraged to help develop novel compounds having suitable pharmacodynamic and pharmacokinetic profiles required of a clinical candidate.