
The Neuroprotective Effect of Activation of Sigma-1 Receptor on Neural Injury by Optic Nerve Crush
By Yao Chen; Xueli Yang; and Junfeng Mao
Excerpt from the article published in Investigative Ophthalmology & Visual Science September 2023, Vol.64, 9. Doi:https://doi.org/10.1167/iovs.64.12.9
Editor’s Highlights
- Sigma-1 receptor (S1R) plays a critical role in mitigating the retinal damage caused by optic nerve crush (ONC).
- S1R protects Retinal ganglion cells (RGCs) against excitotoxic damage by regulating oxidative stress in Müller cells and the secretion of neuroprotective factors through the Nrf2 signaling pathway.
- Selective S1R agonist, resulted in an increase in both basal and maximal mitochondrial respiration, as well as the stabilization of mitochondrial membrane potential.
- S1R potentially facilitates cellular survival by directly inducing the expression of brain-derived neurotrophic factor (BDNF) and that this protective role encompasses the maintenance of retinal structural integrity, promotion of neuronal survival, and safeguarding of visual function.
Abstract
Purpose: This study aimed to explore the neuroprotective effects of sigma-1 receptor (S1R) on optic nerve crush (ONC) mice by upregulating its expression through intravitreal injection of adeno-associated virus (AAV).
Methods: The animals were divided into four groups. Mice that underwent ONC were administered an intravitreal injection with blank vector (ONC group), with AAV targeting downregulation of S1R (S1R-sh group), or with AAV targeting overexpression of S1R (S1R-AAV group). Mice in the control group underwent intravitreal injection with blank vector. The thickness of each layer of the retina was measured through optical coherence tomography, and the apoptotic rate of retinal neurons was determined using the TUNEL assay. The expression levels of brain-derived neurotrophic factor (BDNF) and S1R were quantified through western blot. Electroretinogram (ERG) was performed to evaluate the visual function.
Results: The thickness of the total retina (P = 0.001), ganglion cell layer (P = 0.017), and inner nuclear layer (P = 0.002) in S1R-AAV group was significantly thicker than that of the ONC group. The number of retinal apoptotic cells in the S1R-AAV group was 23% lower than that in the ONC group (P = 0.002). ERG results showed that, compared to the ONC group, the amplitudes of the a- and b-waves were higher in the S1R-AAV group (a-wave, P < 0.001; b-wave, P = 0.007). Western blot showed that the expression of BDNF in the S1R-AAV group was higher than that in the ONC group (P < 0.001).
Conclusions: Activation of S1R in the retina through intravitreal injection of AAV can effectively maintain the retina structure, promote neuronal cell survival, and protect visual function
Introduction
The retina is derived from the central nervous system during the embryonic stage1 and cannot regenerate after injury.2,3 Retinal ganglion cells (RGCs) transmit light from the retina to the superior colliculus and lateral geniculate body through axons.4 They constitute the only nerve conduction pathway and are a potential target for therapies for optic neuropathies.5 Studies have shown that optic nerve crush (ONC) can cause acute optic nerve axon loss, declines in retinal function, and structural changes, particularly damage in the ganglion cell layer and RGC death.6,7
Sigma-1 receptor (S1R) is abundantly expressed in the central nervous system.8 The precise role of S1R in diverse ocular tissues remains incompletely understood; however, the literature indicates that S1R may serve a protective function for ganglion and photoreceptor cells, as evidenced by both in vitro and in vivo experiments. Specifically, numerous reports have demonstrated that these receptors mitigate neuron loss in vitro and contribute to the repair of acute neurodegeneration in vivo.9 S1R expression is enriched in the eye and may inhibit degenerative changes in the eye to promote the survival of lens cells, regulation of intraocular pressure, and retinal neuroprotection of the retina.10 S1R knockout mice showed decreased electroretinogram (ERG) b-wave amplitude, reduced negative scotopic threshold responses, dysfunction of the inner retina (denoted by increased TUNEL-positive cells and active caspase-3–positive cells), and significant loss of the ganglion cells (especially in the central retina) at 12 months.11 The mice also showed damage to the microstructure of optic nerve axons at 6 months of age.12
In Müller cells and ganglion cells, S1R is expressed in the endoplasmic reticulum and nuclear membrane.10 It prevents oxidative stress-induced RGC death in vivo by regulating intracellular calcium ion levels and reducing gene activation before apoptosis. Many studies have confirmed that S1R regulates the response of retinal Müller cells to stress.13 The activation of S1R not only inhibits glutamate excitotoxin-induced development of edema in Müller cells14 but also prevents lipopolysaccharide-induced release of inflammatory factors.15 Wang et al.16 cultured Müller cells from S1R knockout mice and found that S1R regulated cellular reactive oxygen and antioxidant factors. Li et al.17 conducted a study wherein they discovered that the overexpression of the S1R through intravitreal injection of adeno-associated virus 2 (AAV2) facilitated the restoration of RGC function. Additionally, the activation of S1R by pentazocine was observed to decelerate the degeneration of RGC cells and enhance RGC activity subsequent to ONC, thereby providing further evidence of the protective properties associated with S1R. In vivo and in vitro studies have confirmed that activated S1R has protective effects on ganglion cells and photoreceptor cells.18 S1R induces protective effects by reducing peroxide product levels, inhibiting the release of inflammatory factors, and regulating cellular apoptosis.19
Neuronal survival benefits from the persistent inhibition of apoptotic pathways by trophic factors in the microenvironment, and brain-derived neurotrophic factor (BDNF) is crucial in endogenous repair.20 BDNF in the retina is produced endogenously by ganglion cells and glial cells or retrogradely transported from the brain by ganglion axons.21 Studies have found that in patients with glaucoma, the transport of BDNF is blocked, and apoptosis of ganglion cells increases.22 Increased BDNF expression through exogenous supplementation or genetic modification increases ganglion cell survival in ocular hypertension and optic nerve crush models.23,24 At present, S1R agonists have been shown to enhance BDNF expression and downstream signal transduction in retinal astrocytes,25whereas overexpression of S1R promotes precursor BDNF maturation and extracellular secretion.26
In this study, a model of acute optic nerve injury was developed and an intravitreal injection of AAV targeting S1R was administered to investigate the impact of S1R expression on retinal structure, retinal cell apoptosis, and visual function in mice subjected to optic nerve crush. The findings revealed the involvement of BNDF in S1R-mediated neuroprotection and established a novel foundation for the promotion of retinal cell survival following optic nerve injury by S1R.
Results
Construction and Evaluation of the AAV Vector
The AAV interference vector (pAAV-U6-shRNA-CMV-EGFP-WPR), the AAV overexpression vector (pAAV-CMV-EGFP-P2A-SigmaR1-3xFLAG-WPRE), and the blank vector (pAAV-CMV-EGFP-P2A-3xFLAG-WPRE) that were used in this study are shown in Figures 1A, 1B, and 1C, respectively. The results of western blot experiments showed that there was a significant difference in the expression of S1R in the retina of mice after virus injection. Compared with the ONC group, the expression of S1R in the S1R-AAV group was upregulated, whereas in the S1R-sh group the expression of S1R was downregulated (Fig. 1D).
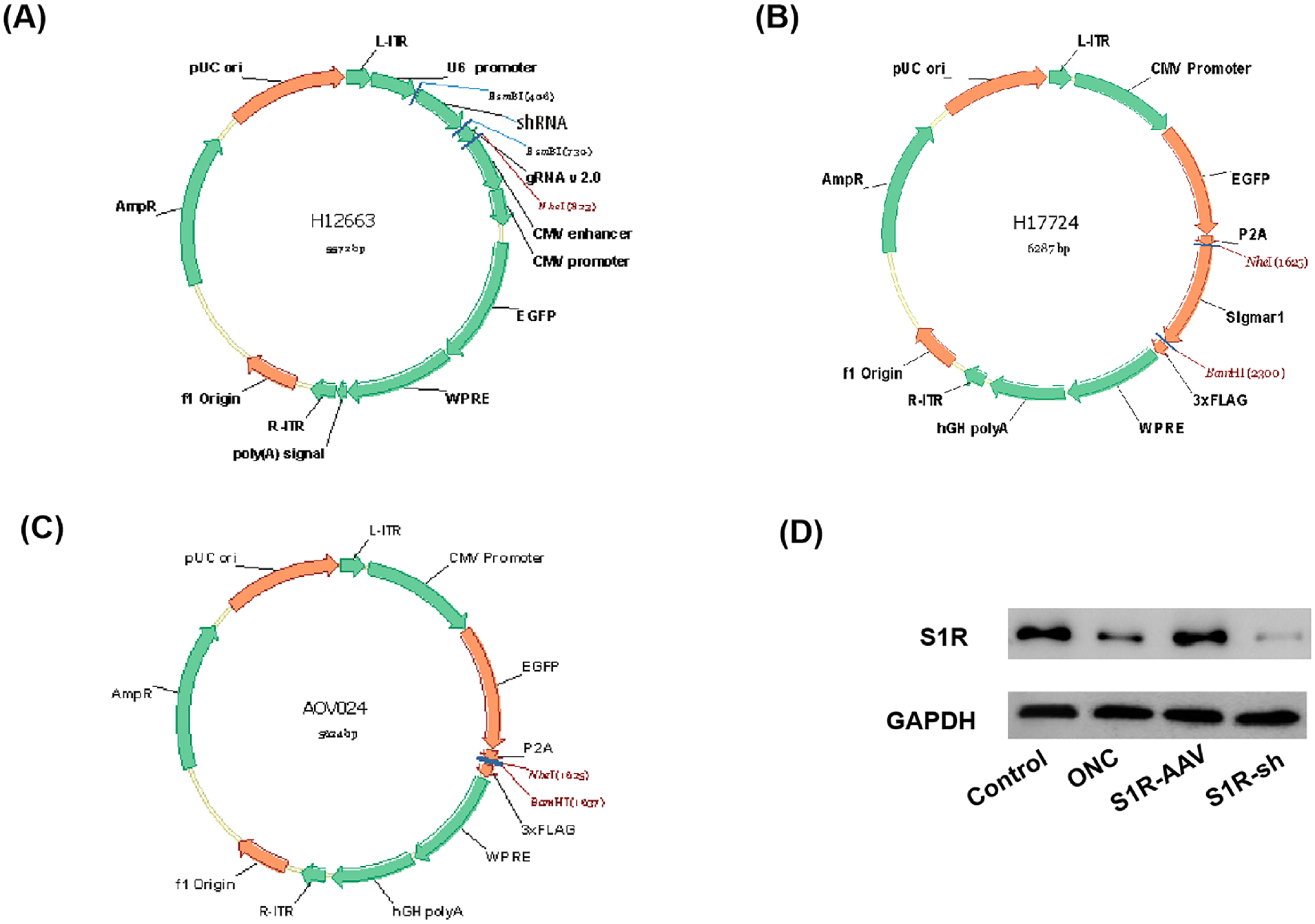
The construction of AAV vectors. (A) AAV interference vector pAAV-U6-shRNA-CMV-EGFP-WPR. (B) AAV overexpression vector pAAV-CMV-EGFP-P2A-SigmaR1-3xFLAG-WPRE. (C) Blank vector pAAV-CMV-EGFP-P2A-3xFLAG-WPRE. (D) Western blot for the expression of S1R in the control group, ONC group, S1R-AAV group, and S1R-sh group.
The vectors carry enhanced GFP so the transfection can be indirectly observed through the degree and location of fluorescence expression. Four weeks after intravitreal injection of AAV, the green fluorescence associated with AAV was distributed in the retina, mainly in the GCL and Müller cells (Fig. 2). The intensity of S1R fluorescence staining was higher in the S1R-AAV group than in the ONC group, and the staining was enhanced in cells located in the GCL and INL. The retina in the S1R-sh group only showed faint red fluorescence (Fig. 2). The above results revealed the effectiveness of the constructed AAV vectors and their capability to integrate into the retina.

Distribution of AAV and S1R expression in mouse retina 4 weeks after AAV transfection. AAV is shown in green, DAPI in blue, and S1R in red.
Expression of S1R in the Eye Maintained the Structure of the Retina
OCT was performed for retinal morphological structure observation and thickness measurement. The morphology of the retina in the control group reveals distinct layers that are arranged in an orderly fashion (Fig. 3A). In the ONC group (Fig. 3B) and S1R-sh group (Fig. 3D), the retinal layers appear smudged and unclear, with distinct retinal thinning in the S1R-sh group. Compared with the ONC and S1R-sh group, the retina in the S1R-AAV group is thicker, and the structure of each layer is relatively ordered and clear (Fig. 3C). Comparing retinal thickness (mean ± SD), the total retinal thickness for the control group was 185.0 ± 3.5 µm; for the ONC group, 145.5 ± 16.7 µm; for the S1R-AAV group, 164.1 ± 10.1 µm; and for the S1R-sh group, 137.6 ± 9.3 µm. The GCL thickness for the control group was 12.8 ± 2.8 µm; for the ONC group, 8.4 ± 1.6 µm; for the S1R-AAV group, 11.2 ± 2.2 µm; and for the S1R-sh group, 6.2 ± 1.9 µm. The INL thickness for the control group was 42.38 ± 3.8 µm; fo the ONC group, 28.8 ± 6.8 µm; for the S1R-AAV group, 35.8 ± 2.6 µm; and for the S1R-sh group, 33.1 ± 3.1 µm. Measurements of retinal thickness in the control group were consistently thicker than those in the ONC group (total retina, P < 0.0001; GCL, P < 0.0001; INL, P < 0.0001), and the S1R-AAV group also showed higher retinal thickness than the ONC group (total retina, P = 0.001; GCL, P = 0.017; INL, P = 0.002). There was no significant difference in retinal thickness between the ONC group and the S1R-sh group. The thickness of the total retina and INL was higher in the control group than in the S1R-AAV group (total retina, P = 0.0002; INL, P = 0.002).
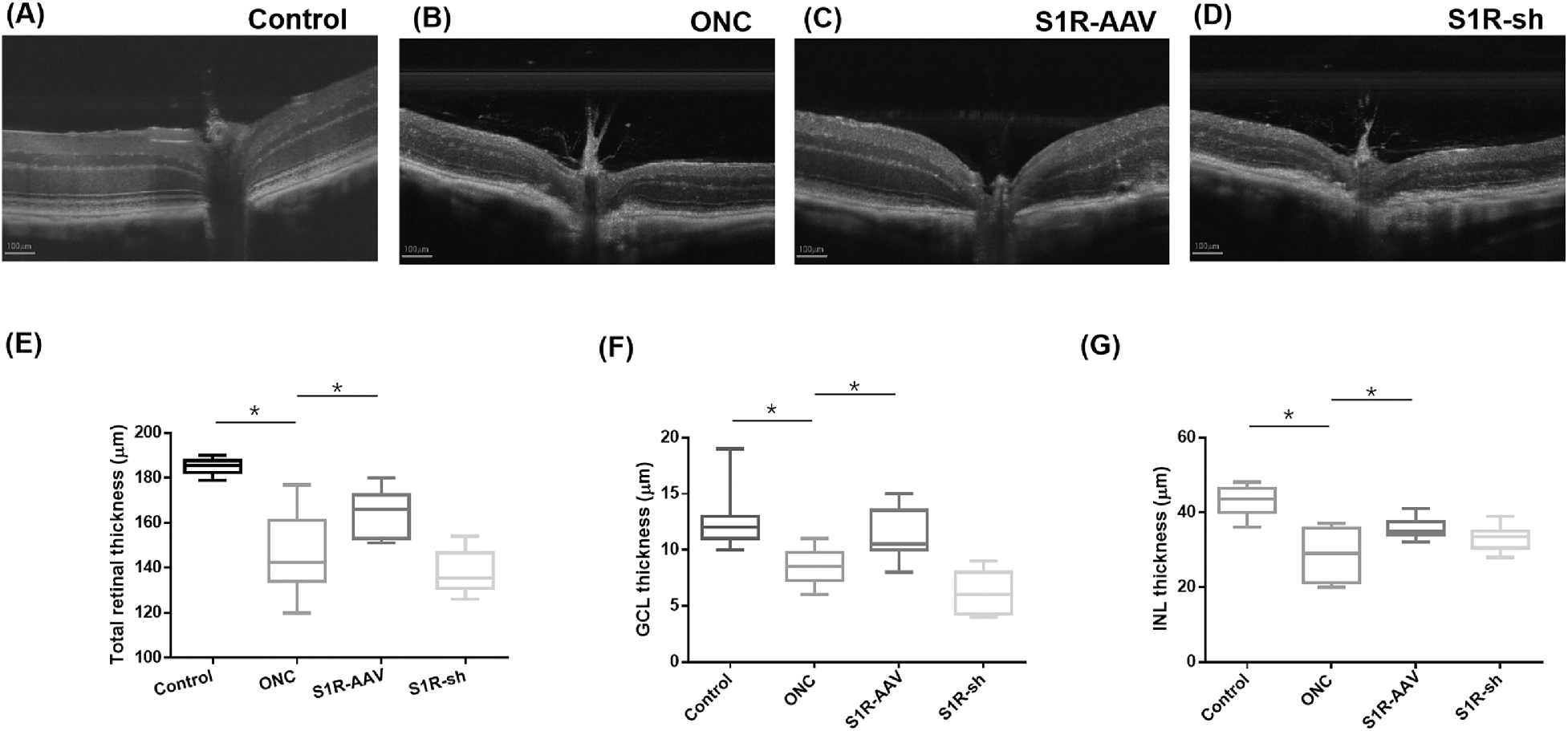
The thicknesses of the retinal layers. (A–D) OCT revealed the retinal thickness of the control group (A), the ONC group (B), the S1R group (C), and the S1R-sh group (D). (E–G) Box plots compare the thickness of retinal layers between groups: (E) total retinal thickness, (F) GCL thickness, and (G) INL thickness. Statistically significant at *P < 0.05.
Activation of S1R Improved Survival of Retinal Cells
Because upregulation of S1R preserved the retinal structure and thickness, we performed H&E staining and TUNEL assays in the control group, ONC group, and S1R-AAV group. H&E staining indicated that the retinal cells in the control group were orderly and tightly arranged (Fig. 4A). In the ONC group, the cells in the INL were relatively loosely arranged, whereas the ganglion cells decreased in number and reduced in size with cytoplasmic vacuolation and pyknotic nuclei (Fig. 4B). Compared with the ONC group, the S1R-AAV group presented improved retinal structure, increased density of ganglion cells and INL nuclei, and alleviated cell degeneration (Fig. 4C). In the TUNEL assay, positive TUNEL staining showed green fluorescence, with blue fluorescence of DAPI showing nuclei. The nuclei with green and blue fluorescence were judged to be apoptotic cells. The positive rate (mean ± SD) in the TUNEL assay in the control group was 0.043 ± 0.024; in the ONC group, 0.284 ± 0.113; and in the S1R-AAV group, 0.056 ± 0.022. The number of apoptotic cells in the ONC group was significantly higher than in the control group (P < 0.001) (Fig. 4D), and the cells were mainly distributed in the GCL, INL, and outer nuclear layer (ONL) (Fig. 4E). Compared with the ONC group, S1R-AAV showed reduced apoptotic cells (P = 0.002) with the TUNEL-positive rate decreased by approximately 23% (Figs. 4F, 5A).
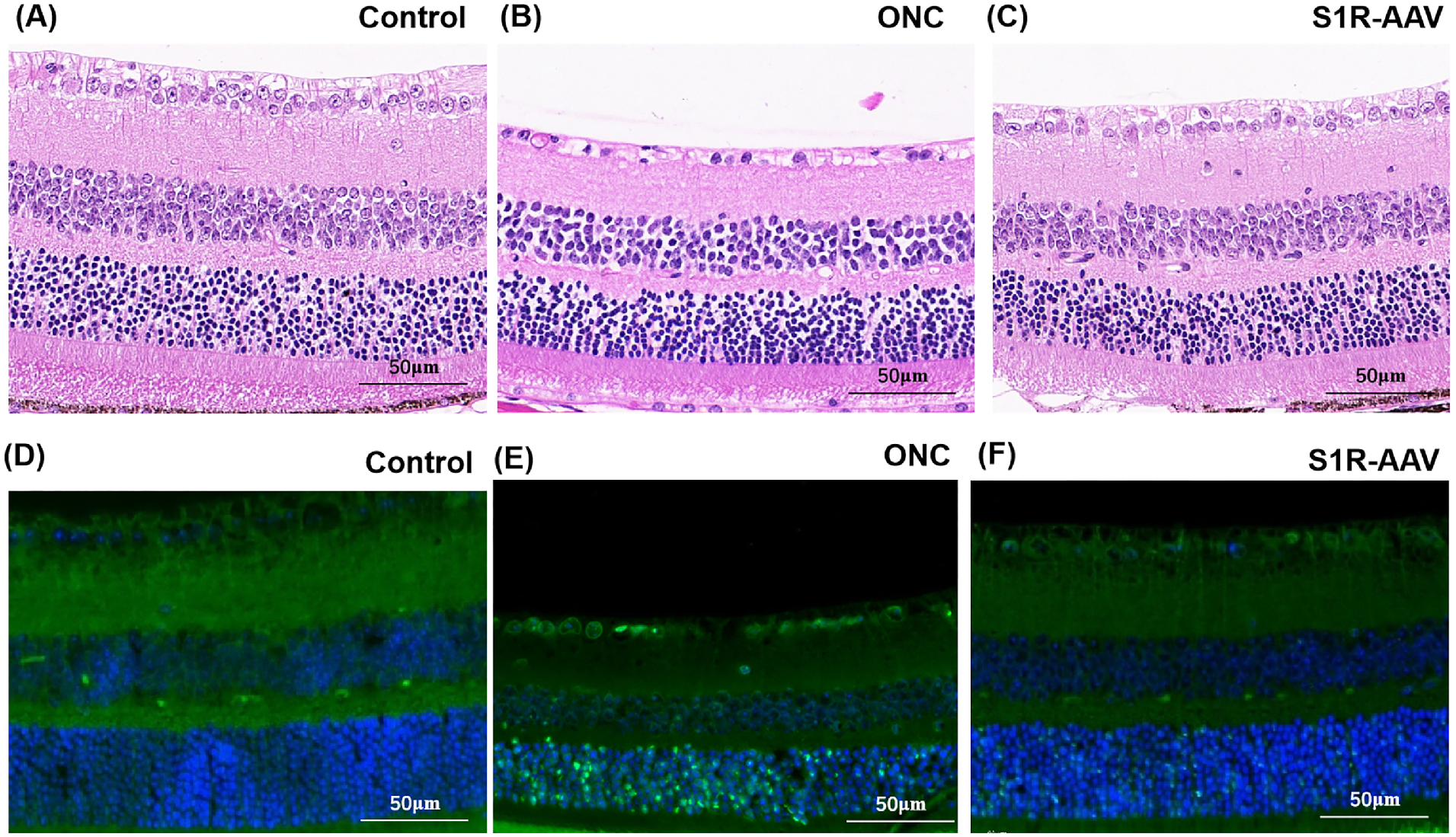
H&E staining and TUNEL assay of the retina. (A–C) H&E staining of retinas in the control group (A), the ONC group (B), and the S1R-AAV group (C). (D–F) TUNEL staining of retinas in the control group (D), the ONC group (E), and the S1R-AAV group (F).
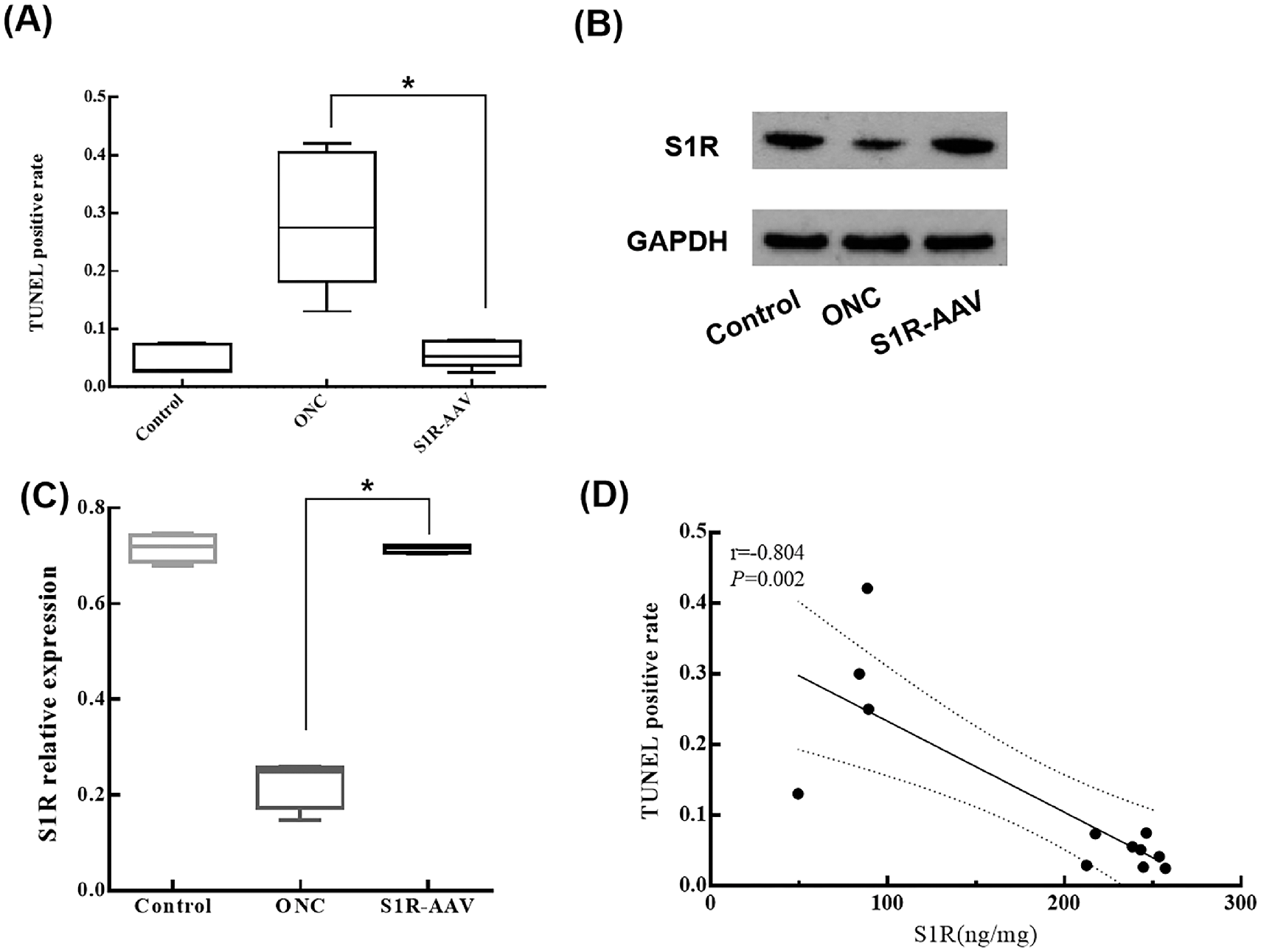
TUNEL-positive rate and expression of S1R. (A) TUNEL-positive rates in the control group, ONC group, and S1R-AAV group. (B) Western blot analyses of S1R of the control group, ONC group, and S1R-AAV group. (C) Relative expression of S1R in the control group, ONC group, and S1R-AAV group. (D) Correlation analysis between the TUNEL-positive rate and S1R expression. Statistically significant at *P < 0.05.
We subsequently performed western blot to understand the effect of S1R on the TUNEL-positive rate (Fig. 5B). Compared with the control group, the relative expression of S1R (the expression ratio of S1R/GAPDH) significantly decreased in the ONC group (P < 0.0001). The relative expression of S1R was elevated in the S1R-AAV group compared to the ONC group (P < 0.0001) (Fig. 5C). Pearson correlation analysis was performed on the expression level of retinal S1R and the TUNEL-positive rate of retinal cells among these three groups, and it showed a negative correlation between S1R and TUNEL-positive rate (r = −0.804, P = 0.002) (Fig. 5D).
S1R Promotes the Expression of BDNF
The results above showed that the activation of S1R preserved the retinal structure and promoted the survival of neural retinal cells. By immunofluorescence, the fluorescent intensity of BDNF was weaker in the ONC group (Fig. 6B) compared with the control group (Fig. 6A) and the S1R-AAV group (Fig. 6C). We verified that the expression of S1R in the S1R-AAV group was higher compared with the control and ONC groups through western blot analysis of the retina (Fig. 6D). The relative expression of BDNF (the expression ratio of BDNF/GAPDH) of the ONC group was lower than that of the control group (P = 0.002) and the S1R-AAV group (P < 0.0001) (Fig. 6E). There was a positive correlation between BDNF and S1R content (r = 0.979, P < 0.001) (Fig. 6F).

The relation of BDNF and S1R expression. (A–D) Immunofluorescence staining of BDNF (red) of the retina in the control group (A), the ONC group (B), and the S1R-AAV group (C). (D) Western blot of retinal BDNF in the control, ONC, and S1R-AAV groups. (E) Relative expression of S1R in the control, ONC, and S1R-AAV groups. (F) Correlation analysis between BDNF and S1R expression. Statistically significant at *P < 0.05.
Activation of S1R Preserved the Function of the Eye
We performed ERG on mice to evaluate the visual function (Figs. 7A–7C). In the control group, the amplitude (mean ± SD) of the a-wave was 199.80 ± 46.19 µV, and the amplitude of the b-wave was 459.80 ± 128.98 µV. In the ONC group, the amplitude of the a-wave was 42.48 ± 19.74 µV, and the amplitude of the b-wave was 121.90 ± 67.42 µV. In the S1R-AAV group, the amplitude of the a-wave was 116.47 ± 13.57uV, and the amplitude of the b-wave was 280.43 ± 66.95 µV (Figs. 7D, 7E). ERG analysis showed that the amplitudes of the a- and b-waves were significantly higher in the S1R group than in the ONC group (a-wave, P = 0.001; b-wave, P = 0.02), but significantly lower than that of the control group (a-wave, P < 0.0001; b-wave, P = 0.002), suggesting that S1R preserved mice visual function in part.

ERG responses and maximum a- and b-wave amplitudes. (A–C) ERG responses of mice in the control group (A), the ONC group (B), and the S1R-AAV group (C). (D) Comparison of ERG maximum a-wave amplitudes among the three groups. (E) Comparison of ERG maximum b-wave amplitudes among the three groups.
Discussion
S1R is widely distributed in the neural retina and is mainly expressed in the endoplasmic reticulum and nuclear membrane of Müller cells and ganglion cells, as well as the nuclear membrane of photoreceptors, bipolar cells, and amacrine cells.12 In this experiment, the AAV was mainly distributed in the GCL and Müller cells after intravitreal injection of AAV. Optic nerve crush–induced retinal damage is mainly manifested in changes in the retinal structure, thinning of the inner neural retina, and apoptosis of GCL and inner nuclear cells.27 The observed reduction in S1R expression within the ocular tissue of mice afflicted with ONC led to significant structural damage to the retina, including disorganized stratification and an unidentifiable GCL. These findings strongly suggest that S1R plays a critical role in mitigating the retinal damage caused by ONC. This is consistent with the results of Ha et al.,11 who observed a significant loss of GCL in S1R–/– mice and increased TUNEL-positive cells. In addition, overexpression of S1R resulted in preserved thickness of the total retina, GCL, and INL and reduced the death of retinal cells by 23%. These findings suggest that the protective effect of S1R was exerted by maintaining retinal structure and the survival of retinal cells. Studies have shown that S1R protects RGCs against excitotoxic damage by regulating oxidative stress in Müller cells and the secretion of neuroprotective factors through the Nrf2 signaling pathway.16 The study conducted by Zhao and colleagues28 demonstrated that the activation of S1R can provide neuroprotection to RGCs in vivo by augmenting the phosphorylation of extracellular signal-regulated kinase 1/2–mitogen-activated protein kinase (ERK1/2/MAPK) under the influence of excitotoxic stress. Similarly, Geva and co-authors29 reported that the administration of pridopidine, a selective S1R agonist, resulted in an increase in both basal and maximal mitochondrial respiration, as well as the stabilization of mitochondrial membrane potential in two distinct rat models of glaucoma, thereby exhibiting neuroprotective properties toward RGCs. Ellis et al.30 demonstrated that the overexpression of S1R mediated by AAV2 in a RGC glucose–oxygen deprivation model significantly augmented the mitochondrial membrane potential while reducing the activity of cytochrome c oxidase and caspase-3/7, thereby enhancing mitochondrial function. Li et al.17 demonstrated that the activation of S1R via intravitreal injection of AAV2 exhibited a mitigating effect on the apoptotic pathway in RGC losses, leading to the restoration of both RGC quantity and functionality. Our experiment revealed that, in addition to preserving RGC, the upregulation of S1R expression in the presence of ONC can also effectively maintain the structural integrity of other retinal layers, including total retinal thickness and INL thickness. The rationale behind this is that S1R has the ability to enhance the viability of retinal cells following damage, as demonstrated by the TUNEL assay. Moreover, the inverse relationship observed between the positive rate of TUNEL and S1R expression provides additional evidence to support this claim. However, future studies are required to determine if direct activation of S1R on Müller cells can promote the repair of neural retina.
To investigate the neuroprotective mechanism of S1R in the context of retinal injury caused by ONC in mice, we conducted an analysis of the expression levels of S1R and BDNF proteins within the retina. BDNF, a potent neuroprotective agent, primarily facilitates the activation of survival effector pathways through its interaction with TrK receptors.31 This interaction enhances downstream signaling, including activation of the PI3K/Akt pathway to inhibit pre-apoptotic targets and activation of the extracellular signal-regulated protein kinase (ERK)/cAMP response element binding protein (CREB) pathway to induce the transcription and expression of genes associated with neuronal survival.32 BDNF exhibits a dual mechanism of action, whereby it not only exerts a direct neuroprotective effect on ganglion cells but also indirectly influences glial cells to upregulate the expression of neurotrophic factors, thereby exerting its effects indirectly.33 Our findings revealed a significant increase in BDNF expression within the S1R-AAV group compared to the ONC group, effectively compensating for the downregulation of BDNF induced by ONC. This suggests a correlation between the upregulation of S1R expression and the observed elevation in BDNF content. In conjunction with the aforementioned experimental findings, it is hypothesized that the S1R potentially facilitates cellular survival by directly inducing the expression of BDNF and that this protective role encompasses the maintenance of retinal structural integrity, promotion of neuronal survival, and safeguarding of visual function in mice.
ERG analysis indicated that S1R activation improved the visual function of mice based on the amplitude of a- and b-waves. It has been reported that S1R–/– mice show decreased ERG b-wave amplitude, decreased negative scotopic threshold responses, and inner retinal dysfunction at 12 months, which is consistent with our results.11 The amplitudes of a- and b-waves were significantly higher in the S1R-AAV group compared with the ONC group. As such, the waveform suggests ameliorated function of rod and cone photoreceptors and bipolar cells. Taken together, the increased expression of S1R improved the visual function of mice after ONC.
To assess the neuroprotective function of S1R, there have been in vivo studies evaluating the effect of activating S1R in alleviating retinal degeneration. Smith et al.34 found that intraperitoneal injection of (+)-pentazocine, a highly specific S1R ligand, in diabetic mice reduced the apoptosis of retinal neurons and effectively alleviated the death of RGC caused by excitotoxins (glutamate and homocysteine) in vitro. In this study, the expression of S1R was upregulated through intravitreal injection of AAV, which is more targeted. AAV has emerged as the most secure viral vector at present due to its minimal immunogenicity and pathogenicity. Furthermore, the augmentation of tissue targeting has enhanced the precision and effectiveness of transfection, rendering it a popular choice for gene transfer to regulate gene expression. In this study, intravitreal viral vector injection was employed to enhance the local tissue virus content and transfection efficiency, resulting in a noteworthy increase in S1R production and endogenous BDNF expression. Through this mode of administration, AAV is more likely to be distributed on the easily damaged GCL and INL, which is more suitable for retinal damages caused by neural degeneration or optic nerve compression.
In conclusion, upregulation of S1R through intravitreal injection of AAV in the retina can effectively maintain the morphology of the retina and protect visual function in mice after ONC. These findings provide deeper insights into the neuroprotective role of S1R and demonstrate the potential of S1R overexpression in the treatment of optic nerve diseases.