
Cocaine Effects on Dopaminergic Transmission Depend on a Balance between Sigma-1 and Sigma-2 Receptor Expression
By David Aguinaga, Mireia Medrano, Ignacio Vega-Quiroga, Katia Gysling, Enric I. Canela, Gemma Navarro, and Rafael Franco
Excerpt from the article published in Frontiers in Molecular Neuroscience, 12 February 2018, Sec. Molecular Signalling and Pathways, Volume 11 – 2018, DOI: https://doi.org/10.3389/fnmol.2018.00017
Editor’s Highlitghts
- In acute cocaine exposure, σ1R modulation of D1R-mediated signaling prevails, but in longer exposures, there is a shift to regulation by σ2R.
- σ2R becomes the main player in conditions of chronic exposure to the drug.
- Some of the addictive and motor actions of cocaine are the result of a balance between cocaine-σ1R versus cocaine-σ2R impact on activation of D1R and D2R.
Abstract
Sigma σ1 and σ2 receptors are targets of cocaine. Despite sharing a similar name, the two receptors are structurally unrelated and their physiological role is unknown. Cocaine increases the level of dopamine, a key neurotransmitter in CNS motor control and reward areas. While the drug also affects dopaminergic signaling by allosteric modulations exerted by σ1R interacting with dopamine D1and D2 receptors, the potential regulation of dopaminergic transmission by σ2R is also unknown. We here demonstrate that σ2R may form heteroreceptor complexes with D1 but not with D2 receptors. Remarkably σ1, σ2, and D1 receptors may form heterotrimers with particular signaling properties. Determination of cAMP levels, MAP kinase activation and label-free assays demonstrate allosteric interactions within the trimer. Importantly, the presence of σ2R induces bias in signal transduction as σ2R ligands increase cAMP signaling whereas reduce MAP kinase activation. These effects, which are opposite to those exerted via σ1R, suggest that the D1 receptor-mediated signaling depends on the degree of trimer formation and the differential balance of sigma receptor and heteroreceptor expression in acute versus chronic cocaine consumption. Although the physiological role is unknown, the heteroreceptor complex formed by σ1, σ2, and D1 receptors arise as relevant to convey the cocaine actions on motor control and reward circuits and as a key factor in acquisition of the addictive habit.
Introduction
In advanced societies cocaine addiction is an important health and socio-economic problem. Cocaine use begins recreationally and the seeking behavior is based on a feeling of general well-being. Drug addiction is the result of plastic changes in areas of the brain that have dopamine as the main neurotransmitter, particularly in the ventral tegmental area(VTA) (see Lüscher, 2013 and references therein). The main consequence of cocaine consumption in the central nervous system (CNS) is an increase in interneuronal dopamine levels, which is not limited to VTA but extends to other structures, such as the basal ganglia (Wise, 1984; Bradberry, 2008). It was thought that the inhibition of dopamine transporters was at the root of all the effects caused by this drug of abuse. However, there is strong evidence showing that cocaine exerts effects by a direct interaction with sigma receptors. Two different sigma receptors have been identified that are functionally and structurally unrelated. Although endogenous ligands are not known and the physiological function of sigma receptors is unclear, these receptors share the ability to bind cocaine. On the one hand, sigma-1 receptor (σ1R) is a chaperone that spans once the membrane bilayer and whose recently reported structure consists of a homotrimer (Schmidt et al., 2016). On the other hand, sigma-2 receptor (σ2R) was identified as a member of the family of membrane-associated progesterone receptors; apart from σ2R (PGRMC1), three other human members are identified: PGRMC2, neuferricin, and neudesin. They are haem proteins displaying a cytochrome b5-fold domain. While σ2R dimerization affects proliferation and chemoresistance in tumor/metastasis in vitro models and xenograft- based tumor/metastasis models, the mode of action in the periphery and the CNS are virtually unknown (Kabe et al., 2016; reviewed in Cahill, 2017). The interaction of σ1R with dopamine receptors and the relevant role that σ1R exerts on the modulation of dopaminergic signaling by cocaine has been reported. In contrast, no study has been undertaken to know whether the binding of cocaine to σ2R results in dopaminergic regulation.
The role of σ1R as relevant target of cocaine was suspected due to the moderate affinity of drug binding to the receptor (Matsumoto et al., 2003; Hayashi and Su, 2005). Therefore, it seems that the “physiologically” relevant concentrations of cocaine can both inhibit the uptake of dopamine and activate σ1R. σ1R-cocaine interaction intervenes in the triggering of locomotor and convulsive actions of the drug (Menkel et al., 1991; Matsumoto et al., 2001a,b, 2002; Barr et al., 2015). In addition, synthetic drugs that act as σ1R agonists and antagonists, respectively, potentiate (Matsumoto et al., 2002, 2003) and reduce (Matsumoto et al., 2004) cocaine actions. More recent studies have identified in both heterologous expression systems and natural sources an interaction between σ1R and dopamine receptors (Navarro et al., 2010; Moreno et al., 2014; Borroto-Escuela et al., 2017). Accordingly, it has been suggested that dual antagonism of σ1R and inhibition of the dopamine DAT transporter can effectively block cocaine self-administration (Katz et al., 2016). Matsumoto et al. (2007) reported that treatment with synthetic drugs that act on σ2R attenuates cocaine-derived behavior in mice. Although the selectivity of the compounds was poor, 1 year later, Mésangeau et al. (2008) designed an approach for converting selective σ1R ligands into σ2R selective ligands that, importantly, showed anti-cocaine activity. Furthermore, it has been observed that treatment with σ2R antagonists counteract locomotor stimulation induced by cocaine in mice (Lever et al., 2014; Guo and Zhen, 2015).
An important physiological consequence of cocaine ingestion is an increase in motor activity, which is controlled by basal-ganglia brain circuits. Motor control is exerted by the direct and indirect pathways of the basal ganglia and associated nuclei. Of the five types of dopamine receptors, the D1 (D1R) is enriched in the direct pathway, while the D2 (D2R) is enriched in the indirect route. The balance of the dopaminergic input in the two circuits results in fine-tuning motor control. The locomotor hyperactivity resulting from cocaine use probably reflects a lack of balance in these two routes. The objective of this work was to investigate how the binding of cocaine to σ2R affects dopaminergic signaling mediated by D1R and/or D2R. We first investigated whether σ2R interacts with D1R or with D2R and, subsequently, we observed how cocaine could affect in a σ1R-independent but σ2R-dependent fashion the signal transduction triggered by agonist activation of D1R but not of D2R.
Results
σ2R May Form Complexes with Dopamine D1 But Not with Dopamine D2Receptors
Two different sigma receptors have been described, the non-opioid receptor, σ1R, and the PGRMC-1 protein, also known as σ2. Despite the endogenous ligands are not known, the two sigma receptors may bind cocaine. While recent studies have demonstrated that σ1R is involved in cocaine modulation of dopamine receptor function, a similar study on σ2R-mediated modulation of dopaminergic signaling is lacking. We first evaluated in a heterologous expression system whether σ2R may colocalize with dopamine receptors at the plasma membrane. Immunocytochemistry assays were undertaken in HEK-293T cells expressing σ2R fused to Rluc and either dopamine D1R fused to YFP or dopamine D2R fused to YFP. The σ2R expression was identified by a specific antibody against Rluc protein and a secondary Cy3 antibody, while dopamine receptor-YFP expression was identified by its own fluorescence. D1R (green) was detectable at the plasma membrane level while σ2R (red) was expressed both in intracellular structures and at the plasma membrane, where it colocalized (yellow) with D1R (Figure 1A, left images). When a similar experiment was developed with D2R, similar results were obtained indicating that D2R and σ2R colocalize at the cell surface (Figure 1A, right images). When the immunocytochemical assays were performed in cells pretreated with 30 μM cocaine for 30 min, the level of colocalization between σ2R and D1R or D2R was similar, indicating that cocaine pretreatment did affect neither cell surface expression of σ2R, D1R or D2R nor receptor colocalization. Next, we determined whether σ2R may form heteromer complexes with dopamine D1 or D2 receptors. For this purpose, we took advantage of energy transfer assays and in situ proximity ligation assay (PLA), which allows the identification of close proximity between two proteins (<17 nm) (Borroto-Escuela et al., 2011; Trifilieff et al., 2011). For PLA, HEK-293T cells expressing σ2R and either D1R or D2R were treated with specific primary antibodies against σ2R and against each of the dopamine receptors. Interestingly, the red punctuated signal around Hoechst-stained nuclei was much higher for D1R and σ2R than for D2R and σ2R (82 versus 27% of labeled cells) (Figure 1B). Finally, we developed bioluminescence energy transfer assays in HEK-293T cells transfected with cDNAs for D1R-Rluc or D2R-Rluc and increasing amounts of cDNA for σ2R-YFP. Interestingly, a saturable BRET curve was obtained (BRETmax 50 ± 3, BRET50 190 ± 40) (Figure 2A) indicating a specific interaction between D1R-σ2R; in contrast, a linear signal was obtained between D2R-σ2R (Figure 2B) suggesting a lack of interaction between them. When the same experiments were undertaken in cells treated with cocaine, similar results were obtained for the D1R-Rluc/σ2R-YFP donor/acceptor pair (BRETmax 82 ± 10, BRET50680 ± 200), indicating that cocaine did not significantly affect the interaction (Figure 2B).
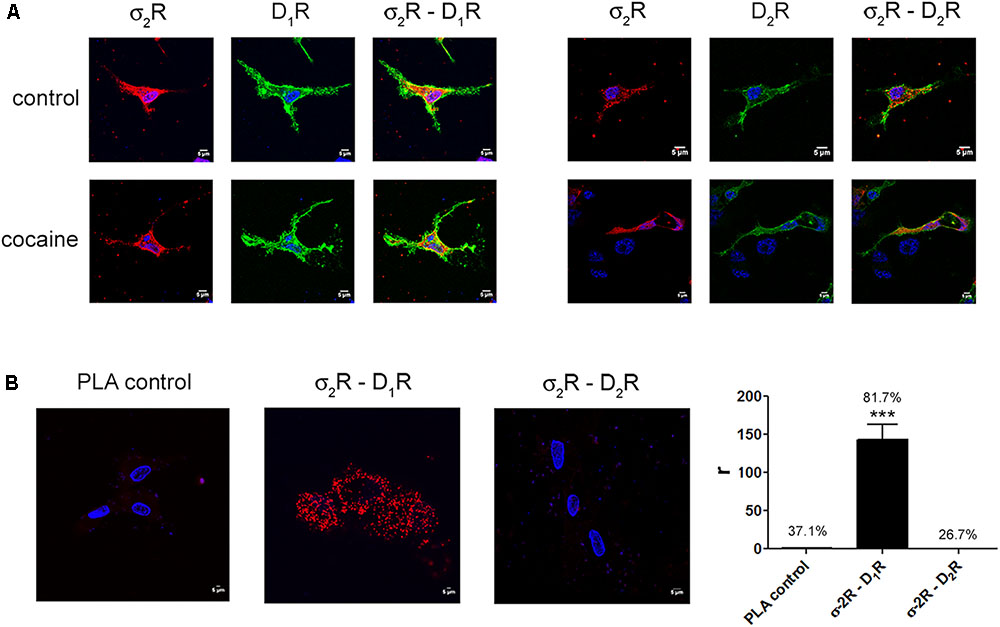
Expression of σ2R-containing heteromer complexes in a heterologous expression system.
To determine colocalization between σ2R and dopamine D1 or D2 receptors, immunocytochemistry assays were performed in HEK-293T pretreated or not with 30 μM cocaine for 30 min. HEK-293T cells expressing σ2R-Rluc (1 μg cDNA), D1R-YFP (1 μg cDNA), D2R-YFP (1 μg cDNA), σ2R-Rluc (1 μg cDNA) and D1R-YFP (1 μg cDNA) or σ2R-Rluc (1 μg cDNA) and D2R-YFP (1 μg cDNA) were used. Dopamine receptors were detected by YFP fluorescence (green) and σ2R was detected by a specific antibody against Rluc (1/100, Millipore, Temecula, CA, United States) followed by a Cy3-conjugated secondary antibody (1/200, Jackson Immunoresearch Laboratories, West Grove, PA, United States) (red). Colocalization is shown in yellow (A). Scale bar 5 μm. In situ proximity ligation assay (PLA) was developed in HEK-293T cells expressing D1R (1 μg cDNA) or D2R (1 μg cDNA) and σ2R (1 μg cDNA) by the use of specific primary antibodies (1/100 dilution) against D1R, D2R and/or σ2R. Nuclei were stained with Hoechst (1/100). Confocal microscopy images (four superimposed sections) were obtained showing D1R-σ2R or D2R-σ2R complexes as red spots (B). Scale bar 5 μm. Quantification of the PLA provides in the Y-axis the ratio r (number of red spots/cell containing spots) and, above each bar, the percentage of positive cells versus the total number of cells (blue nucleus). Data are the mean ± SEM of four different fields in five independent preparations. One way ANOVA and Dunnett’s post hoc test showed statistically significant differences (∗∗∗p < 0.001).
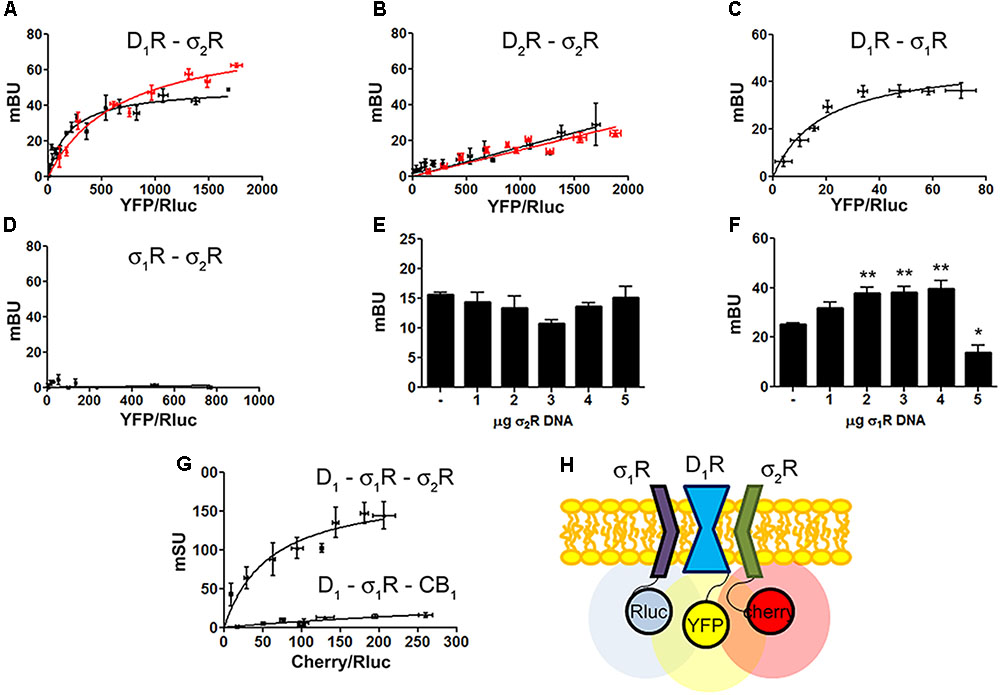
Identification of D1R-σ1R-σ2R heteromers in a heterologous expression system.
Bioluminescence energy transfer (BRET) was developed in HEK-293T cells expressing (i) a constant amount of D1R-Rluc (0.3 μg cDNA) (A)or D2R-Rluc (0.25 μg cDNA) (B) and increasing amounts of σ2R-YFP (0.5–4 μg cDNA), or (ii) a constant amount of D1R-Rluc (0.3 μg cDNA) and increasing amounts (0.5–4.5 μg cDNA) of σ1R-YFP (C) or (iii) a constant amount of σ1R-Rluc (0.1 μg cDNA) and increasing amounts of σ2R-YFP (1–6 μg cDNA) (D). Cells were treated (red line) or not (black line) with 30 μM cocaine for 30 min. BRET is expressed as milli BRET units (mBU) and is given as the mean ± SEM of seven different experiments. Competition experiments were developed in HEK-293T cells expressing a constant amount of σ1R-Rluc (0.1 μg cDNA) and D1-YFP (1.5 μg cDNA) and increasing amounts of unfused σ2R (0–5 μg cDNA) (E) or a constant amount of D1-Rluc (0.05 μg cDNA) and σ2R-YFP (0.3 μg cDNA) and increasing amounts of unfused σ1R (0–5 μg cDNA) (F). Transfer of energy was expressed as milli BRET units (mBU) and results are given as the mean ± SEM of 10 different experiments. One way ANOVA and Dunnett’s post hoc test showed statistically significant differences (∗p < 0.05, ∗∗p < 0.01). (G) Sequential resonance energy transfer (SRET) assay developed in HEK-293T cells transfected with constant amounts of σ1R-Rluc (0.2 μg cDNA) and D1R-YFP (1.5 μg cDNA) and increasing amounts of σ2R-RFP (0.5–4 μg cDNA). A negative control was performed using cDNA for the cannabinoid CB1receptor (fused to RFP) instead of cDNA for σ2R-RFP. SRET is expressed as milli SRET units (mSU) and are given as the mean ± SEM of 6 different experiments. (H) Schematic representation of SRET.
Dopamine D1R, σ1R, and σ2R May Form Heterotrimeric Complexes
Dopamine D1 and σ1 receptors may form heteromeric complexes in HEK-293T cells (Navarro et al., 2010). To confirm whether in our experimental conditions D1R-Rluc may act as a donor of σ1R-YFP, BRET experiments undertaken in cotransfected HEK-293T cells provided a saturable curve thus indicating the interaction between σ1R and D1R (Figure 2C). We then hypothesized that σ1R and σ2R could be interacting together. Accordingly, BRET assays were performed in HEK-293T cells expressing a constant amount of σ1R-Rluc and increasing amounts of σ2R-YFP. The unspecific linear signal obtained (Figure 2D) suggested that no interaction was occurring between the two sigma receptors. We then performed assays to investigate whether σ1R and σ2R competed for the binding to D1R. BRET experiments were then developed in HEK-293T cells expressing a constant amount of σ1R-Rluc and D1-YFP and increasing amounts of non-fused σ2R. The results indicated that σ2R was not able to compete with σ1R for heteromer formation since the energy transfer between donor and acceptor was not altered (Figure 2E). When a similar experiment was performed expressing a constant amount of D1-Rluc and of σ2R-YFP and increasing amounts of non-fused σ1R, the results indicated that low expression levels of σ1R increased BRET signals; however, higher expression levels σ1R were able to displace σ2R out of the heteromer, as reflected by a significant decrease in BRET signal (Figure 2F). This result could reflect the formation of D1R-σ1R-σ2R heterotrimer complexes, where the interaction of σ1R to the σ2R-D1R complex could create a structural change in turn leading to increasing the energy transfer between Rluc and YFP. To confirm this possibility, sequential resonance energy transfer (SRET) assays, which permits detection of trimers (Carriba et al., 2008), were developed in HEK-293T cells expressing a constant amount of σ1R-Rluc and of D1R-YFP and increasing amounts of σ2R-Cherry. The saturable SRET curve indicates that formation of σ1R-D1R-σ2R heteromer complexes was occurring (Figures 2G,H). The negative control was performed by substituting σ2R-RFP by the cannabinoid CB1 receptor fused to RFP thus confirming the specificity of the triple σ1R-D1R-σ2R interaction (Figure 2G).
σ2R Activation Blocks Dopamine D1R Signaling
Our next aim was to characterize the functionality of the σ1R-D1R-σ2R heterotrimer structure in HEK-293T cells treated with cocaine. It should be noted that σ1R (Navarro et al., 2010) and σ2R (Johannessen et al., 2011) are endogenously expressed in HEK-293T cells; consequently, we used a siRNA approach to silence σ1R or σ2R expression thus impeding heterotrimer formation. When HEK-293T cells were transfected with D1R and siRNA for σ1R, SKF-81297-induced a significant increase in cAMP levels, that was inhibited by pretreatment with cocaine or with the σ2R agonist, PB-28, indicating that cocaine decreases D1R-mediated cAMP signaling function through its binding to σ2R (Figure 3A). When HEK-293T cells were transfected with D1R and siRNA for σ2R, the results indicated that cocaine pretreatment potentiated agonist-induced cAMP levels, which was evidence of cocaine action upon binding to the σ1R (Figure 3B). The next set of results is consistent with a reciprocal modulation of signaling mediated by cocaine binding to σ2R and σ1R; while cocaine via σ1R positively modulates cAMP levels, it inhibits cAMP signaling via σ2R. Accordingly, no effect of cocaine was observed in HEK-293T expressing D1R and the two endogenous sigma receptors (Figure 3C). The lack of modulation exerted by cocaine upon simultaneous binding to both σ1R and σ2R likely reflects a balance which would, in a physiological set-up, depend on the relative expression of the two sigma receptors. In fact, when HEK-293T cells were transfected with D1R and both siRNA for σ1R and σ2R, cocaine or the specific σ2R agonist, PB-28, had no effect, indicating that cocaine modulation over D1R depends on σ1R and σ2R expression (Figure 3D).
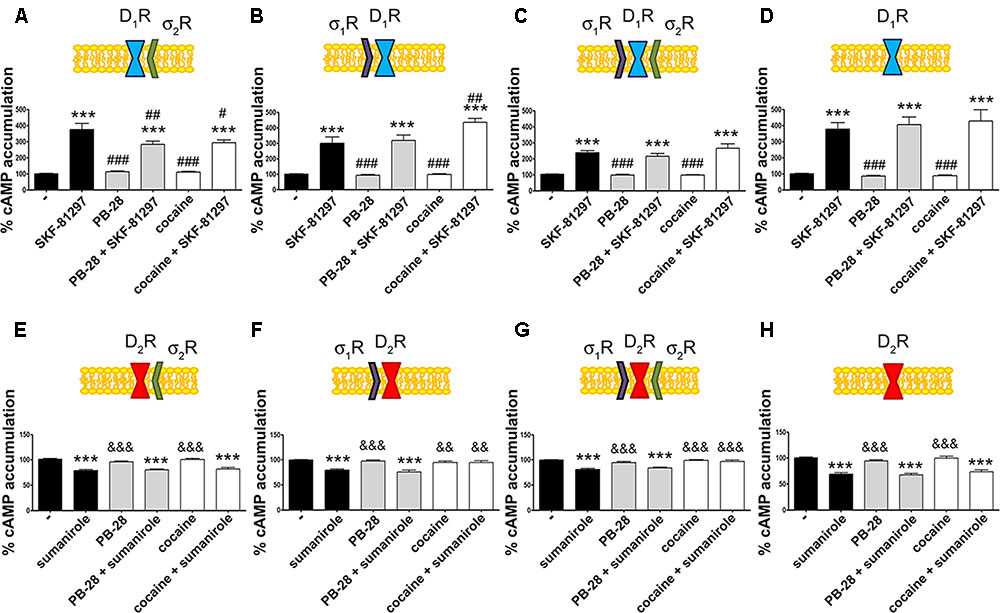
σ2R modulation of D1 receptor-mediated signaling in a heterologous expression system.
cAMP determination experiments were developed in HEK-293T cells expressing D1R (A–D) or D2R (E–H) in the absence (C,G) or presence of 3 μg siRNA for σ1R (A,E), 3 μg siRNA for σ2R (B,F) or both (D,H). Cells were pretreated with 30 μM cocaine for 30 min, 300 nM PB-28 or vehicle 15 min prior to receptor activation using 200 nM SKF-81297 or 500 nM sumanirole. In cells expressing D2R 0.5 μM forskolin was used to induce increases in cAMP levels. Basal [cAMP] is considered 100% in cells expressing D1R, whereas forskolin-induced [cAMP] is considered 100% in cells expressing D2R. Values are the mean ± SEM of 12–15 different experiments. One way ANOVA followed by a Dunnett’s multiple comparison post hoc test showed a significant effect of treatments versus control (∗∗∗p < 0.001), a significant effect of treatments versus SKF-81297 (#p < 0.05, ##p < 0.01, and ###p < 0.001) and a significant effect of treatments versus sumanirole (&&p < 0.01 and &&&p < 0.001).
We next investigated whether cocaine binding to σ2R receptors could still modulate D2R-mediated signaling. HEK-293T cells transfected with cDNAs for D2R and siRNA for σ1R, responded to the selective-D2R agonist, sumanirole. In these cells the Gi-mediated decrease of forskolin-induced cAMP accumulation due to Gi coupling was not affected by cocaine pretreatment (Figure 3E). These results agree with the lack of interaction between σ2R and dopamine D2R (see Figure 2B). As a control, we confirmed that when the σ1R-cocaine modulation over D2R was assayed, i.e., silencing σ2R expression, cocaine was able to block the sumanirole-induced effect (Figure 3F). These results agree with those in Navarro et al. (2013) in the sense that they reflect the consequence of a physical interaction between σ1R and D2R receptors. In agreement with this hypothesis, HEK-293T cells expressing D2R and endogenous sigma receptors behaved as cells in which the σ2R was silenced (Figure 3G). As a further control, HEK-293T cells treated with siRNAs to silence both sigma receptors showed no modulation by cocaine over D2R-mediated signaling (Figure 3H), thus reinforcing the idea that cocaine effect over D2R depends on σ1R expression.
σ2R Activation Potentiates Dopamine D1R MAP Kinase Phosphorylation
To further understand the cocaine effect over D1R function, MAP kinase signaling was evaluated in HEK-293T cells transfected with cDNAs for D1R and siRNA for either σ1R or σ2R. In cells expressing D1R with silenced σ1R, i.e., expressing D1R and σ2R (Figure 4A), cocaine pretreatment increased agonist (SFK-81297)-induced ERK1/2 phosphorylation, while in cells with silenced σ2R, i.e., expressing D1R and σ1R (Figure 4B), cocaine decreased agonist-induced ERK1/2 phosphorylation. These results are evidence of potentiation by cocaine-σ2R of MAP kinase signaling, and potentiation by cocaine-σ1R of Gs-protein dependent signaling. In cells expressing D1R and the two sigma receptors, no effect of cocaine pretreatment on pERK1/2 levels was observed, in agreement with the above-described balance resulting from reciprocal sigma-receptor-mediated cocaine effects (Figure 4C). As a further control, cocaine did not alter the SKF-81297-induced ERK1/2 phosphorylation in HEK-293T cells expressing D1R and with silenced sigma receptors (Figure 4D). A similar experimental design was used to undertake dynamic mass redistribution (DMR) assays. DMR is a label-free technique useful to investigate the activation of G-protein coupled receptors (Grundmann and Kostenis, 2015; Medrano et al., 2017). On the one hand, in cells expressing D1R and σ2R, cocaine blocked SKF-81297-induced increase in the DMR signal in a similar way as the selective σ2R ligand, PB-28, did (Figure 4E). On the other hand, the SKF-81297 effect was potentiated by cocaine pretreatment in cells expressing D1R and σ1R (Figure 4F). Once more, cocaine modulation on D1R-agonist-induced effects was not found in cells expressing D1R and both sigma receptors (Figure 4G). As DMR in cells expressing D1R mainly reflects Gs-coupling (Kebig et al., 2009; Schröder et al., 2009; Hamamoto et al., 2015), these results are similar to those obtained in cAMP read-outs. Another control was performed to show that pretreatment with the σ2R selective agonist, PB-28, did not result in any signal modulation in cells expressing D1R but silenced σ1R and σ2R expression (Figure 4H).
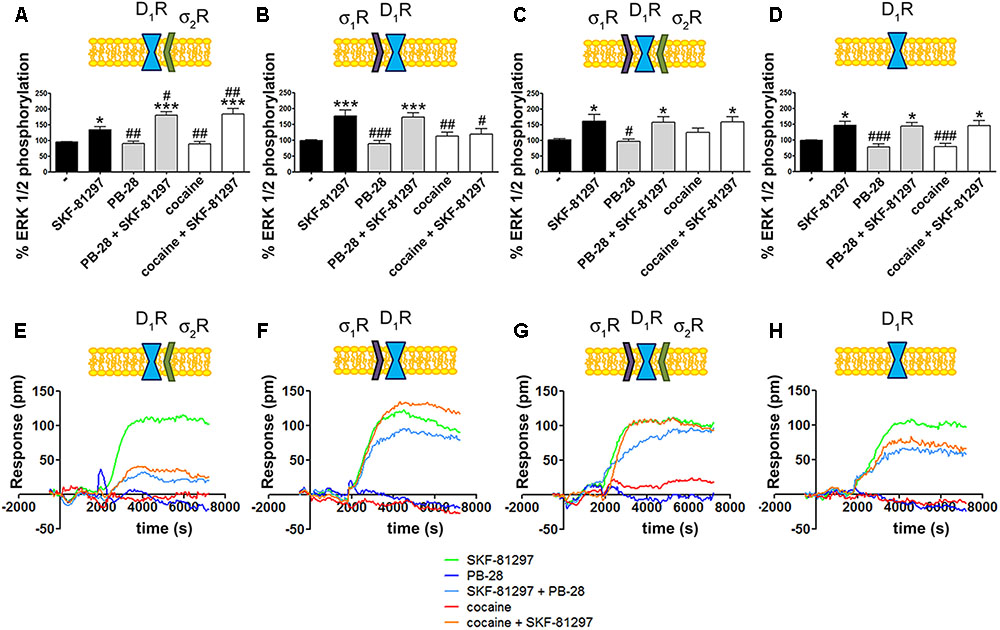
Cocaine effects on D1R-mediated signaling.
MAP kinase activation was determined in HEK-293T cells transfected with 0.75 μg cDNA for D1R in the absence (C) or presence of 3 μg siRNA for σ1R (A), 3 μg siRNA for σ2R (B) or both (D). The culture medium was replaced by non-supplemented DMEM and 2 h later cells were treated for 30 min with 30 μM cocaine, 300 nM PB-28 or vehicle followed by a 200 nM SKF-81297 stimulation (7 min). The basal level of pERK1/2 is considered 100%. Values are the mean ± SEM of 10–12 different experiments. One way ANOVA followed by a Dunnett’s multiple comparison post hoc test showed a significant effect of treatments versus control (∗p < 0.05, ∗∗∗p< 0.01) and a significant effect of treatments versus SKF-81297 (#p < 0.05, ##p < 0.01, and ###p< 0.001). Real-time DMR signal 60 min recordings in HEK-293T cells transfected with 0.75 μg cDNA for D1R in the absence (G) or presence of 3 μg siRNA for σ1R (E), 3 μg siRNA for σ2R (F) or both (H) that were treated with 30 μM cocaine (red), 300 nM PB-28 (dark blue) or vehicle (green) for 30 min previous to 200 nM SKF-81297 stimulation.
σ2R Activation Blocks Dopamine D1R-Mediated Signaling in Primary Cultures of Striatal Neurons
A proximity ligation assay (PLA) was used to determine in primary cultures of striatal neurons whether D1R-σ2R complex expression was affected by cocaine pretreatment. Consequently, specific antibodies against D1R and σ2R were used in neurons treated or not with cocaine for 30 min (Figure 5A). 32% of cells showed punctuated staining (with 2.2 red spots/cell containing spots) surrounding Hoechst-stained nuclei (Figure 5B). These results indicate the occurrence of D1-σ2 heteroreceptor complexes in striatal primary cultures of neurons. A control done in the absence of primary antibodies led to 18% of labeled cells (with 1.2 red spots/cell containing spots). The percentage of positive cells after a 30-min treatment with cocaine was around 30 (with 2 red spots/cell containing spots) (Figure 5B). Thus, cocaine pretreatment did not significantly alter D1R-σ2R complex formation. When PLA was developed to detect D2R and σ2R complexes, the results (19% with 1.3 red spots/cell containing spots) were similar to those in the negative control (20% with 1.4 red spots/ cell containing spots), i.e., no evidence of heteroreceptor formation was obtained. Pretreatment with cocaine did not lead to the appearance of heteromer complexes formed by D2R and σ2R (Figure 5B). These results agree with the BRET assays that did not find sign of interaction between the D2R-Rluc and σ2R-YFP but between the D1R-Rluc and σ2R-YFP pair.
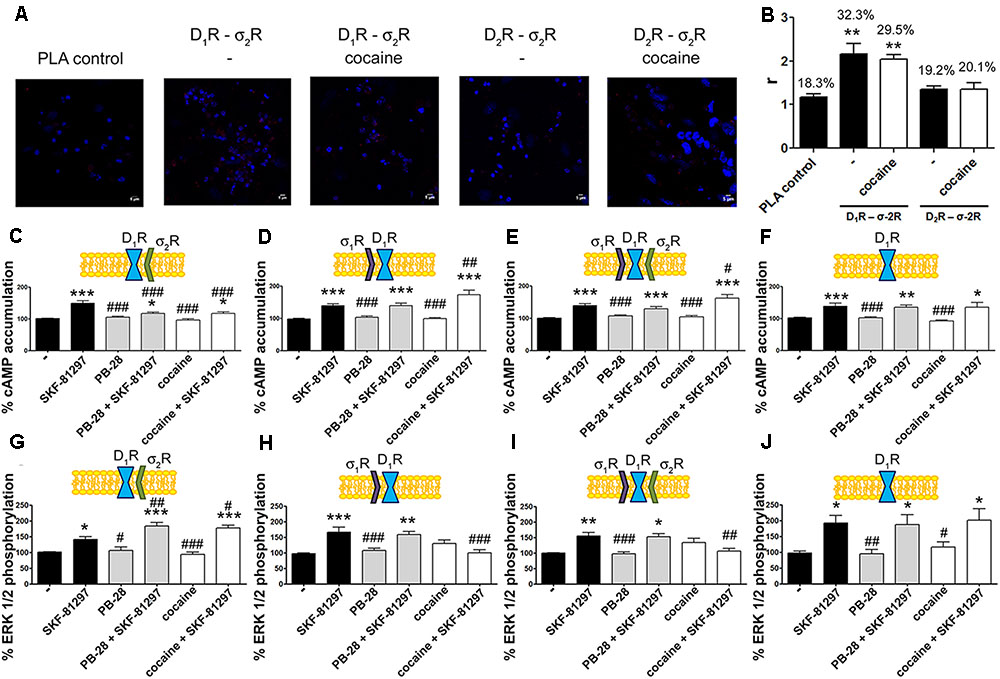
Expression and function of σ2R-D1R complexes in primary cultures of striatal neurons.
In (A,B) PLA assay was developed in striatal primary cultures of neurons pretreated or not with cocaine 30 μM for 30 min. σ2R-D1R or σ2R-D2R heteromer complexes were detected by the use of specific antibodies (1/100 dilution) against σ2R and D1R or σ2R and D2R. Confocal microscopy images (four superimposed sections) were obtained where nuclei were stained with Hoechst (1/100). Scale bar 5 μm (A). Quantification of the PLA provides in the Y-axis the ratio r (number of red spots/cell containing spots) and, above each bar, the percentage of positive cells versus the total number of cells (blue nucleus) (B). Data are the mean ± SEM of four different fields in five independent preparations. One way ANOVA and Dunnett’s multiple comparison post hoc test showed statistically significant differences versus control (∗∗p < 0.01). Primary cultures of striatal neurons, control (E,I) or transfected with siRNA for σ1R (C,G), σ2R (D,H) or both (F,J) were treated with 30 μM cocaine for 30 min or 300 nM PB-28 prior to 200 nM SKF-81297 stimulation. cAMP levels (C–F) or MAP kinase activation l (G–J) were determined. Basal [cAMP] is considered 100%. The basal level of pERK1/2 is considered 100%. Values are the mean ± SEM of 10–15 different experiments. One way ANOVA followed by a Dunnett’s multiple comparison post hoc test showed a significant effect of treatments versus not treated cells (∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001) and a significant effect of treatments versus SKF-81297 (#p < 0.05, ##p < 0.01, and ###p< 0.001).
To demonstrate the effect of cocaine over D1R-mediated signaling in a more physiological environment, we analyzed cAMP and MAP kinase signaling pathways in primary cultures of striatal neurons. As striatal neurons express the two sigma receptors, the siRNA approach was used to silence sigma receptor expression. On the one hand, in neurons transfected with siRNA for σ1R, and consequently expressing D1R and σ2R, cocaine and PB-28 led to a decrease in agonist-induced cAMP levels and to an enhancement in MAP kinase signaling (Figures 5C,G). On the other hand, in neurons transfected with the siRNA for σ2R, and consequently expressing D1R and σ1R, cocaine but not PB-28 induced an increase in the cAMP signal and a decrease in the ERK1/2 phosphorylation signal (Figures 5D,H). Most of these results agree with those obtained in the heterologous system. However, in striatal neurons expressing D1R and both sigma receptors, cocaine treatment led to a net effect that showed predominance of σ1R- versus σ2R-mediated modulation (Figures 5E,I). These findings could be due to a higher expression of σ1R-D1R complexes versus σ2R-D1R but they may also result from the lower affinity of the cocaine/σ2R binding (Lever et al., 2016). Finally, another control was performed to show that pretreatment with the σ2R selective agonist, PB-28, or with cocaine, did not result in any signal modulation in cells expressing D1R but silenced σ1R and σ2R expression (Figures 5F,J).
D1R-Mediated Signaling Is Modulated by σ1R in Acute and by σ2R in Chronic Conditions
In situ PLAs were performed to identify D1R-σ1R and D1R-σ2R heteroreceptor complexes in striatal sections from Sprague–Dawley rats receiving cocaine under acute or chronic regimes (see section “Materials and Methods”) (Figure 6A). When striatal sections of vehicle-treated animals were analyzed, it was observed that 38.5% of cells showed D1R-σ1R complexes with 2.5 red spots/cell containing spots, while only 25% of cells showed D1R-σ2R complexes with 2.1 dots/cell (Figure 6B). When Sprague–Dawley rats were acutely treated with cocaine, it was observed that both D1R-σ1R and D1R-σ2R complex expression increased. However, the D1R-σ1R complexes doubled its expression while D1R-σ2R complex expression suffered a slight increase (respectively, 54% of cells showed red spots with 4.5 spots/cell and 33% with 2.3 spots/cell). Interestingly, in the case of rats chronically treated with cocaine, the D1R-σ1R heteromer complex expression was not affected (34% of cells showed red spots with 2.4 spots/cell containing spots) compared to control animals, while the D1R-σ2R heteromer expression significantly increased (35% of cells containing spots with 3.4 spots/cell containing spots) (Figure 6B). These results indicate that acute cocaine treatment strongly increases D1R-σ1R complexes formation in striatal rat sections but chronic cocaine treatment only drives D1R-σ2R complex expression. Then, we questioned if the cocaine-induced alterations in D1R-σ1R and D1R-σ2R complex expression had signaling consequences. To do so, we analyzed SKF-81297-induced cAMP production in primary cultures of striatal neurons pretreated with vehicle or cocaine for different times (from 0.5 h to 7 days). Interestingly, we observed that at short times SKF-81297-induced cAMP levels were further increased. In agreement with results in HEK-293 cells, cocaine binding to the σ1R induced a positive modulation over dopamine D1R-mediated signaling. When primary cultures of neurons were longer exposed to cocaine (1–7 days), SKF-81297-induced increase in cAMP levels was inhibited (Figure 6C). Taking into account the results in HEK-293 cells such effect seems associated to D1R-σ2R complex formation and to the ability of σ2R to counteract the SKF-81297-induced increases of cAMP. To check whether these interpretations were correct, i.e., if sigma receptors were responsible of cocaine-induced modulations over D1R-mediated signaling, primary striatal neurons were transfected with siRNA specific for σ1R or σ2R. On the one hand, cocaine pretreatment (0.5 h to 7 days) blocked SKF-81297-induced accumulation of cAMP levels in primary cultures of neurons transfected with siRNA for σ1R, i.e., expressing D1R and σ2R (Figure 6D). On the other hand, 0.5 h and 2 h pretreatment of cocaine potentiated the SKF-81297-induced increases in cAMP levels in primary neurons transfected with siRNA for σ2R, i.e., expressing D1R and σ1R. However, longer periods of cocaine exposure (1–7 days) produced no effect (Figure 6E). These results suggest that in acute cocaine treatment D1R form heteromers mainly with σ1R, prevailing the D1R-σ1R-mediated signaling. In contrast, in the chronic situation, the increase of σ1R-D1 heteromer complex expression observed in acute conditions disappear but the increase in the D1R-σ2R complex expression is maintained, being the σ2R responsible of the cocaine modulation over D1R, hence prevailing the D1R-σ2R-mediated signaling.
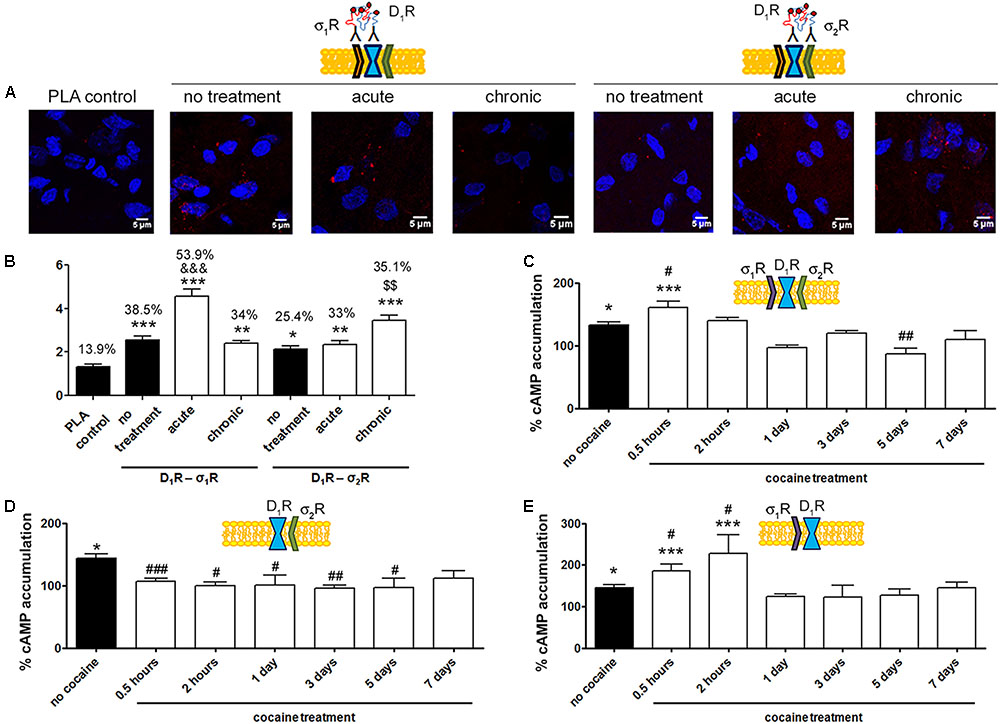
D1R-mediated signaling is modulated by σ1R in cocaine acute and by σ2R in cocaine chronic exposure.
In (A,B) PLA assay was developed in brain sections from male Sprague–Dawley rats i.p. injected with vehicle or 15 mg/kg cocaine under acute or chronic regimes (see section “Materials and Methods”). D1R-σ1R or D1R-σ2R heteromer complexes were detected by PLA using of specific antibodies against D1R (1/100), σ1R (1/100, Santa Cruz Biotechnology, Dallas, TX, United States) or σ2R (1/100). Confocal microscopy images (four superimposed sections) were obtained where nuclei were stained with Hoechst (1/100). Scale bar 5 μm (A). Quantification of the PLA provides in the Y-axis the ratio r (number of red spots/cell containing spots) and, above each bar, the percentage of positive cells versus the total number of cells (blue nucleus) (B). Data are the mean ± SEM of six different fields in five independent preparations. One way ANOVA and Dunnett’s multiple comparison post hoc test showed statistically significant differences versus control (∗p < 0.05, ∗∗p < 0.01, ∗∗∗p < 0.001), significant differences in D1R-σ1R complex amount between acute and chronic treatments (&&&p < 0.001) and significant differences in D1R-σ2R heteromer complex amount between acute and chronic treatments ($$p < 0.01). cAMP determination experiments were developed in primary cultures of striatal neurons, control (C), transfected with 3 μg siRNA for σ1R (D) or 3 μg siRNA for σ2R (E). Cultures were divided into 9 groups and pretreated with vehicle or 30 μM cocaine for different time periods (from 0.5 h to 7 days) prior to receptor activation using 200 nM SKF-81297. Basal [cAMP] is considered 100%. Values are the mean ± SEM of five different experiments. One way ANOVA followed by a Dunnett’s multiple comparison post hoc test showed a significant effect of treatments versus basal (∗p < 0.05, ∗∗∗p < 0.001) and a significant effect of cocaine treatments (white bars) versus cocaine non-exposed neurons (black bar) (#p < 0.05, ##p < 0.01, and ###p < 0.001).
Discussion
Sigma receptors are relevant in cocaine addiction, because binding of cocaine to these receptors modulates dopaminergic transmission. Although cocaine can bind to both σ1R and σ2R, they are not closely related and no common structural properties have been identified. With respect to the modulation of receptor-mediated signaling, a relevant difference is revealed by the formation of heteroreceptor complexes. On the one hand, σ1R interacts with D1 and D2 dopamine receptors (Navarro et al., 2010; Moreno et al., 2014). Interestingly, we here report that σ2R may form heteromeric complexes with D1R but not with D2R.
Information on PGRMC1/σ2R expression in brain is partial. Intlekofer and Petersen (2011)confirmed data by Krebs et al. (2000) showing enrichment of the receptor in nuclei of the hypothalamus that are important for female reproduction. Petersen et al. (2013) in 2013, reviewed neuroanatomical data on the expression PGRMC1 and related proteins in CNS neuroendocrine nuclei. To our knowledge, information on expression in other neural regions is either absent or preliminary. Interestingly, a recently developed fluorescent probe tested in rat brain indicates that the receptor is more present in neurons than in glial cells (Zeng et al., 2016). Despite good in vitro properties, some of the radiolabeled probes that were developed for in vivo σ2 receptor imaging have not reached the final objective (Abate et al., 2013; Selivanova et al., 2015). In contrast, recently reported 18F-labeled PET probes, with enhanced brain uptake and σ2R selectivity (in mice), show promise for in vivo imaging of the receptor in the human brain (Wang et al., 2017). Surely these novel tools will be instrumental to achieve a more detailed mapping of the receptor in the CNS, specially in those areas in which dopamine receptors are expressed. There is, however, strong evidence of expression in the striatum; for instance, a recent report shows that receptor agonist regulate dopaminergic input into the striatum and the receptor is presynaptically expressed the nucleus accumbens (Klawonn et al., 2017). Furthermore, pioneering studies by Werling and colleagues showed σ2R involvement in control of dopamine transporter activity in striatum (Derbez et al., 2002) and that the striatal receptor was a target of cocaine (Nuwayhid and Werling, 2006).
The results presented here and those already reported (Navarro et al., 2010, 2013) show that in equivalent experimental configurations, cocaine binding to σ1R improves the accumulation of cAMP mediated by D1R and inhibits MAP kinase signaling. Cocaine, via σ2R, blocks D1-mediated cAMP accumulation and enhances MAP kinase activation. Importantly, similar results were obtained in HEK-293T cells and primary neuronal cultures.
D1R can form complexes and high- order heteromers by interacting simultaneously with the σ1 and σ2 receptors, σ1R being able to displace σ2R, but not vice versa. Navarro et al. (2010)reported an increase in the plasma membrane expression of σ1R after acute exposure to cocaine. When increase in σ1R levels in the plasma membrane occurs, σ2R is displaced from the D1R-σ2R or D1R-σ2R-σ1R heteroreceptor complexes. Such phenomenon results in increasing the amount of D1-σ1 heteroreceptors and D1R signaling whereas, as reported by Navarro et al. (2013), reducing D2R-mediated actions. However, in a longer exposure to cocaine, the signaling mediated by the dopamine D1R fits more with that occurring via a D1R-σ2R functional unit. These data suggest that the initial cocaine-induced overexpression in the plasma membrane of the σ1R is transient; once these levels decrease, due to internalization or other still unknown mechanisms, σ2 is the predominant receptor forming heteromers with dopamine D1R.
Motor control in the basal ganglia is achieved through a complex circuit composed of GABAergic neurons that contain mainly D1R (direct pathway) and GABAergic neurons containing mostly D2R (indirect pathway) (Grillner and Robertson, 2016). Fine motor control is achieved by a balance of dopaminergic signals, one via D1 receptors, which are Gscoupled, and another via D2 receptors, which are Gi coupled (Jenner, 1995; Gerfen, 2000). The deterioration of motor control by cocaine depends on the imbalance of the direct/indirect pathway, but the underlying mechanism remains unclear. Although the scenario is complex, cocaine is known to increase cAMP levels in cells expressing D1R-σ1R (Navarro et al., 2010). Therefore, cocaine seems to be increasing in the direct pathway the cAMP-dependent dopaminergic output, namely activation of protein kinase A and cAMP-regulated DARPP-32 phosphoprotein (Svenningsson et al., 2004). Through the same receptor (σ1R), cocaine leads to a deterioration of the dopaminergic performance of the indirect route (Navarro et al., 2013). In addition to the imbalance resulting from these σ1R-dependent effects, our results demonstrate that trimers of D1, σ1, and σ2 receptors may be formed and that cocaine acting on these heteromers reduces the negative modulation exerted by the D1R-σ1R complexes.
The results here presented also show that the MAP kinase signaling pathway is particularly affected by the action of cocaine upon dopamine-sigma heteroreceptors. While in cells expressing the D1R-σ1R heteromer, cocaine decreased ERK1/2 phosphorylation, cocaine did the opposite in cells expressing the D1R-σ2R heteromer. It is known that ERKs are involved in the plastic changes induced by the consumption of drugs of abuse (Radwanska et al., 2005). In addition, the inhibition of ERK phosphorylation alters learned place-preference in a paradigm of drug-of-abuse consumption, whereas activation of ERK1/2 is necessary to establish the association between place preference and drug consumption (Valjent et al., 2006; Du et al., 2017). In this context, knocking down ERK1 has shown that enhanced ERK2 signaling and repeated exposure to the drug facilitate the plastic changes leading to drug addiction (Ferguson et al., 2006). It should be noted that the temporal pattern of MAP kinase activation in the mouse brain is differently induced by addictive or non-addictive drugs (Valjent et al., 2004). Interestingly, Zhang et al. (2017) have described that D1 receptor antagonists alter in cocaine-treated mice the length of nucleus accumbens postsynaptic densities, i.e., cocaine-induced long-term plasticity; however, the mechanism underlying this phenomenon has not been described. According to the previous reports and to our results, it may be suggested that potentiation of MAP kinase pathway mediated by the D1R-σ2R heteromer may be the mechanism by which the σ2R would induce long term neuronal plasticity. The predominant role of the σ1R in acute cocaine use shifts to a more relevant role of σ2R in the chronic condition leading to the establishment of addiction. In any case, the relative expression of the two receptors in a given neuron seems important in determining the fate of the cell when the drug of abuse is consumed.
In acute cocaine exposure, σ1R modulation of D1R-mediated signaling prevails, but in longer exposures, there is a shift to regulation by σ2R. Recently, Singer et al. (2017) have determined that neuronal plasticity initiate 2 h after cocaine exposure. The mechanism of action described in this paper cannot explain some of the results reported by Matsumoto et al. (2007) and Lever et al. (2014) who report that σ2R receptor antagonists block the effects of cocaine-induced hyperlocomotion. It should be, however, noted that a recent report show benefits of a σ2R -selective agonist, siramesine, for decreasing cocaine effects via reduction of dopaminergic and glutamatergic input to the striatum (Klawonn et al., 2017). On the one hand, it is a reasonable assumption that σ1R is more involved in the regulation of D1R signaling at acute exposure. However, it remains to be determined whether some of the results reported on the impact of cocaine on locomotion are due to the use of non-selective ligands, that is, ligands that can bind to both sigma receptors and alter their function. Alternatively, it may happen that σ2R is also affecting the direct route in acute conditions. What our results undoubtedly indicate is that σ2R becomes the main player in conditions of chronic exposure to the drug. In summary, some of the addictive and motor actions of cocaine are the result of a balance between cocaine-σ1R versus cocaine-σ2R impact on activation of D1R and D2R (and D1R-D2R, see Perreault et al., 2016) in ad hoc CNS circuits.