
Targeting Sigma-1 Receptor: A Promising Strategy in the Treatment of Parkinson’s Disease

By Talha Siddiqui and Lokesh Kumar Bhatt
Excerpt from the article published in Neurochem Research. 31 May 2023. DOI: https://doi.org/10.1007/s11064-023-03960-6
Editor’s Highlights
- Activation of Sigma-1 receptors (S1Rs) by agonists provides neuroprotection via different mechanisms.
- S1R KO mice have shown increased pathogenic alpha-synuclein aggregation and S1R agonists have shown increased degradation of alpha-synuclein.
- S1R activation decreases microglial activation and inflammatory response.
- S1R agonists antagonize NMDA receptors thus inhibiting glutamate-mediated excitotoxicity.
- S1R agonists modulate gene expression of endogenous antioxidants.
- All these properties make S1R an ideal therapeutic target for Parkinson’s disease (PD).
Abstract
Parkinson’s disease is a neurodegenerative disease affecting mainly the elderly population. It is characterized by the loss of dopaminergic neurons of the substantia nigra pars compacta region. Parkinson’s disease patients exhibit motor symptoms like tremors, rigidity, bradykinesia/hypokinesia, and non-motor symptoms like depression, cognitive decline, delusion, and pain. Major pathophysiological factors which contribute to neuron loss include excess/misfolded alpha-synuclein aggregates, microglial cell-mediated neuroinflammation, excitotoxicity, oxidative stress, and defective mitochondrial function. Sigma-1 receptors are molecular chaperones located at mitochondria-associated ER membrane. Their activation (by endogenous ligands or agonists) has shown neuroprotective and neurorestorative effects in various diseases. This review discusses the roles of activated Sig-1 receptors in modulating various pathophysiological features of Parkinson’s disease like alpha-synuclein aggregates, neuroinflammation, excitotoxicity, and oxidative stress.
Introduction
Parkinson’s disease (PD) is one of the most common neurodegenerative diseases. The prevalence of PD is approximately 1–4% of the population over 60–80 years of age [1]. Statistics say more than ten million people have been diagnosed with PD and the numbers are still soaring [2]. The pathognomonic feature in PD post-mortem brains is Lewi bodies since these cytoplasmic inclusions are formed by the aggregation of the alpha-synuclein. Alpha-synuclein is a regulatory protein that controls many neuronal functions including neurotransmitter release [3, 4]. In PD, the gene encoding alpha-synuclein gets mutated and hence causes its aggregation. This depletes the neuronal dopamine level resulting in symptoms like tremors, rigidity, changes in gait, and bradykinesia/hypokinesia. Moreover, the pathogenic alpha-synuclein aggregate promotes neurodegeneration [3]. The post-mortem studies of the brains of PD patients show inflammation in the basal ganglia region [5, 6]. Further, overactivation of NMDA (N-methyl-D-aspartate) receptors causes excitotoicity which causes neuronal apoptosis. The exact cause of PD is still a matter of debate but various pieces of evidence have pointed to oxidative stress and genetics as etiological factors [7, 8]. The conventional pharmacological options for PD provide symptomatic relief mainly. Thus, an unmet need exists to bring new disease-modifying strategies for the treatment of PD.
The main pathophysiological factors associated with PD are excess/misfolded alpha-synuclein aggregates, microglial cell-mediated neuroinflammation, excitotoxicity, oxidative stress, and defective mitochondrial function. Disruption of mitochondrial structure and function is one of the main mechanisms in the pathogenesis of PD. Mitochondria-associated endoplasmic reticulum membrane (MAM) of neurons of the substantia nigra region have ubiquitous expression of Sigma-1 receptors (S1Rs). The S1Rs are molecular chaperones and are associated with multiple functions. S1Rs promote cell survival by activation of the defence mechanisms. S1R activation (by endogenous ligands or agonists) has shown neuroprotective and neurorestorative effects in various diseases [9,10,11].
Structure and Function of S1R
Sigma receptors are intracellular proteins expressed throughout the body including the central nervous system. Initially, sigma receptors were defined as a subclass of opioid receptors however later studies confirmed that sigma receptors are non-opioid receptors [12, 13]. Sigma receptors have two types viz. Sigma-1 and Sigma-2. The former has a prominent role in neurodegenerative diseases whereas the latter showed importance in cancer. S1Rs are intracellular chaperones with a molecular weight 29-kDa and total 223 amino acids. It does not have homology with any other mammalian protein except the yeast sterol isomerase ERG2, where it shares only 60 percent of the amino acid sequence. The amino acid sequence identity is 30.3 percent [14]. S1R resides on MAM and is neither an ionotropic receptor nor a metabotropic receptor. This 29-kDa protein has two hydrophobic domains at N-terminus and at C-terminus, it has domains for molecular chaperone activity. The centre of S1R has membrane-spanning α-helices [15,16,17,18]. The N-terminus and C-terminus lie on the same side with N-terminus located at cytosol and C-terminus located at the endoplasmic reticulum. The gene that encodes S1R i.e. SIGMAR1 is ~7 kbp long situated on chromosome 9, band p13. It has 4 exons and 3 introns with exon 3 being the shortest (93 bp) and exon 4 being the longest (1132 bp) [19,20,21]. S1Rs perform crucial roles in the cell like modulation of endoplasmic reticulum (ER) stress, mitochondrial function, glial activity, oxidative stress, and regulation of Ca+ influx. Activation of S1Rs (by endogenous ligands or agonists) decreases misfolded protein-induced ER stress, neuroinflammation, excitotoxicity, and oxidative stress. Thus, S1R poses itself as a promising target and S1R agonist, a valuable extension in pharmacological classes of anti-parkinsonian agents [22,23,24,25].
Role of S1Rs in Managing Proteome
The maintenance of cellular proteins at an optimum level is of utmost importance to govern cell survival. It has been observed that excess or misfolded alpha-synuclein aggregates disturb cellular physiology and contribute to neuronal degeneration in PD [26]. Rectifying the faulty conformation or enhancing the degradation or suppressing the genesis of alpha-synuclein can be a promising strategy to tackle PD. S1Rs being molecular chaperones, control the cellular protein concentration. Excess/misfolded protein induces ER stress which activates two programs viz. unfolded protein response (UPR) and ER overload response (EOR). UPR has 3 sensor proteins (i) IRE1 (inositol-requiring enzyme 1) (ii) PERK (protein kinase RNA-activated-like endoplasmic reticulum kinase) (iii) ATF6 (activating transcription factor 6) which works to bring down ER stress [15, 22, 24, 27,28,29,30].
IRE1 undergoes dimerization and phosphorylation to get converted into its active state. Activated IRE1 splices XBP1 (X-box binding protein 1) mRNA alters the transcription of UPR genes and result in upregulation of ER chaperones. In addition, upregulation of ER chaperones occurs when ATF6 gets activated. These ER chaperones then perform ER-associated protein degradation (ERAD). The activation of PERK reduces global protein translation by inhibiting eIF-2α (eukaryotic translation initiation factor 2 α). PERK activation can also trigger apoptosis. Sustained PERK activation upregulates CHOP (C/EBP-homologous protein) which then inhibits the expression of Bcl-2 (anti-apoptotic factor) and upregulates pro-apoptotic BH3-only proteins expression. This leads to the activation of Bak- and Bax-dependent apoptosis [24]. The S1R aids in modulating the activities of IRE1, PERK, and ATF6 to a significant extent. ER stress, S1R ligand or agonist stimulation dissociates S1R from BiP (binding immunoglobulin protein) and stabilizes IRE1 (Fig. 1). S1R remains dormant when attached to BiP and performs maximum when dissociated. During ER stress, there is upregulation of S1R [15, 22, 24, 31, 32]. S1R agonism can help in alpha-synuclein clearance and prevent neurodegeneration in PD. With regards to EOR, it has been observed that its activation enhances reactive oxygen species (ROS) formation and NF-κB (nuclear factor kappa B) activation [15].
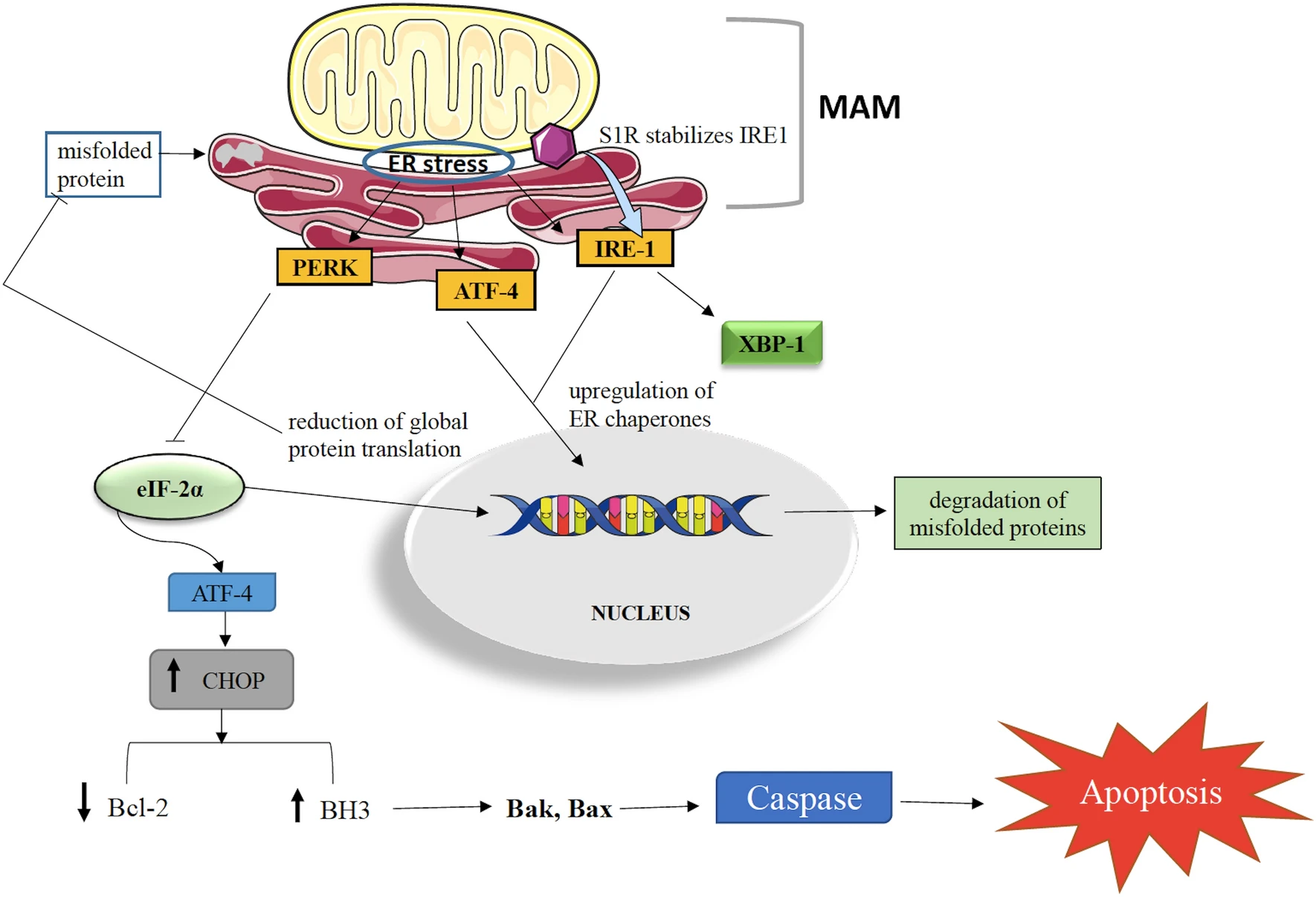
Modulation of misfolded protein-induced ER stress by sigma 1 receptor
Role of S1Rs in Modulating Neuroinflammation
Microglia are macrophage-derived immune cells of the central nervous system (CNS) and constitute 10% of the total cellular population of CNS [33]. Under sound physiological conditions, microglial cells are responsible for many functions like surveillance, neurogenesis, cognition, pathogen destruction, etc. [34]. However, in a pathological state like PD, microglial cells cause neuroinflammation and contribute to neurodegeneration. Further, neurodegeneration leads to neuroinflammation [35]. So far, two phenotypes of microglia have been discovered viz. M1 (classic) and M2 (alternative) [36]. The former is pro-inflammatory and detrimental to CNS whereas the latter is anti-inflammatory and neuroprotective. In PD, M1-type microglial activation releases nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, ROS, and pro-inflammatory cytokines (interleukin-1β,6 and tumor necrosis factor-α) which leads to neuroinflammation [37, 38]. Contrastingly, activation of M2-type microglia decreases pro-inflammatory cytokine levels and performs neuronal repair and regeneration [24, 39, 40].
S1Rs are also expressed on microglial cells and modulate microglial activities. Francardo et al. has demonstrated that S1R activation through PRE-084 and pridopidine reduces CD-68 positive microglial cells in the striatum and substantia nigra region of C57Bl/6 mice. In addition, there was a significant enhancement in TH+ cells suggesting neuroprotective properties of S1R agonists-PRE-084 and pridopidine. Further, the activity of PRE-084 and pridopidine was lost in S1R KO mice, confirming the agonist’s activity via S1R [41, 42]. Afobazole, a clinically approved anxiolytic in Russia, was reported to have affinity towards S1R. Cuevas et al. studied the effects of afobazole in ischemic stroke and found that S1R activation by afobazole decreases microglial migration and inflammatory response. For microglia to activate and migrate, the influx of cations (Ca2+, Na2+) through purinergic receptors (P2X and P2Y), which are localized on microglia, is needed. Activation of these receptors is governed by ATP (ligand of P2X and P2Y). Activated S1R is found to block these receptors and hence possess anti-inflammatory properties (Fig. 2) [33, 43]. Hall et al. reported that 1,3-Di-o-tolylguanidine (a potent S1R agonist) suppresses microglial migration, response, and morphology in a murine model of stroke. Their study suggests that S1R activation regulates various aspects of microglial function [44]. The probable mechanism could be the enhancement of anti-inflammatory cytokines i.e. interleukin-10 and 4 by S1R agonist. These anti-inflammatory cytokines inhibit inflammatory cytokines and hence prevent neuroinflammation (Fig. 2) [9, 45]. Another sigma ligand-SR-31747 was found to inhibit tumor necrosis factor-α (TNF-α) and interferon-γ (IFN-γ) secretion evoked by lipopolysaccharide (LPS) in rats [46]. Similar results were obtained with the study of Wu et al., but they used a different S1R ligand i.e. SKF89359 [47]. All the aforementioned data indicates the anti-inflammatory properties of S1R ligands/agonists.
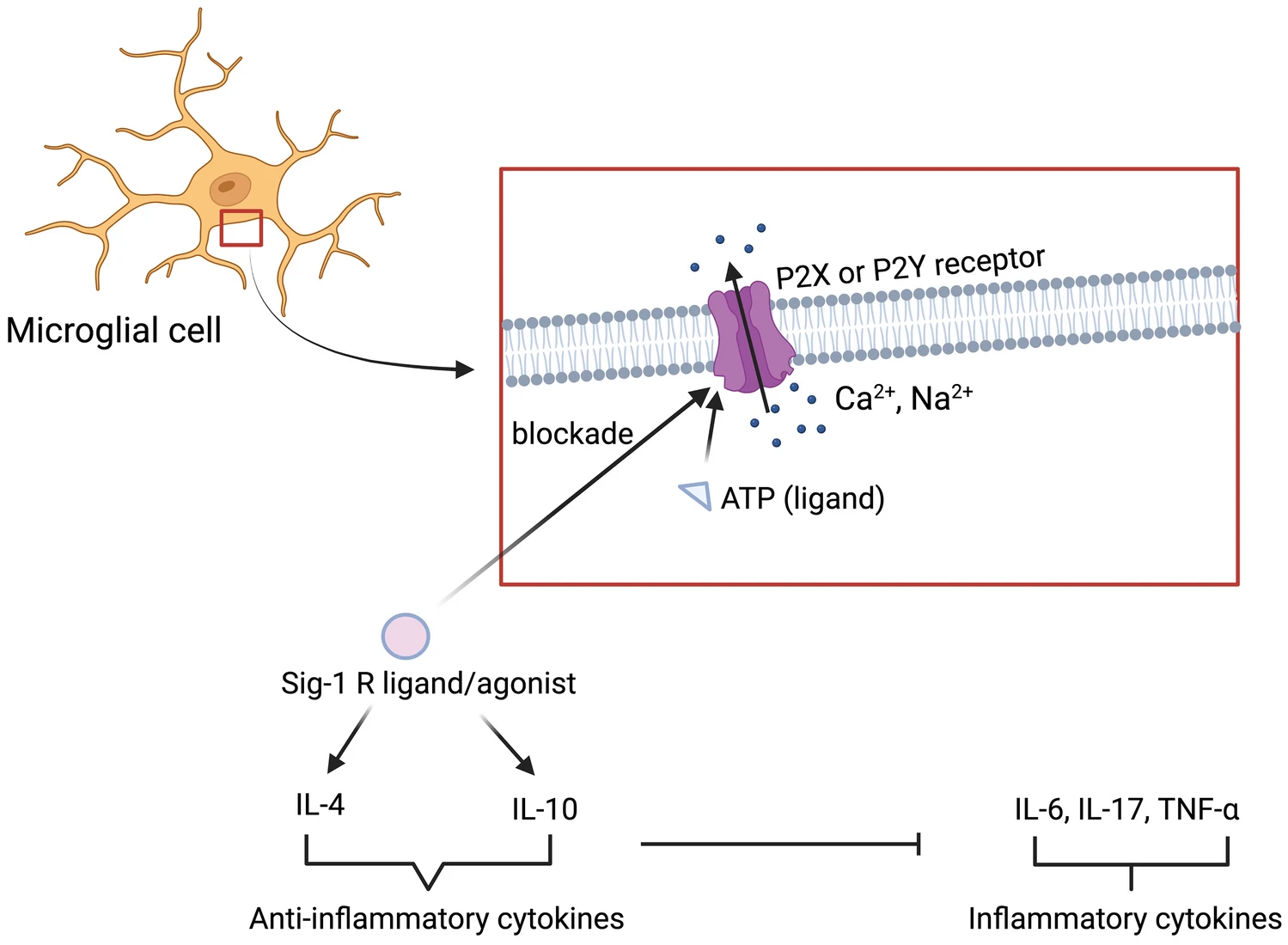
Modulation of neuroinflammation by activation of sigma 1 receptor
Role of S1R in Modulating Excitotoxicity
In the brain, the balance between excitatory and inhibitory neural signals is crucial for healthy neuronal functioning and viability. The excitatory signals are mediated by Glutamate-a major excitatory neurotransmitter. The arrival of action potential/signal stimulates the synaptic vesicle to release glutamate into the synaptic cleft. The glutamate then binds to glutamatergic receptors (majorly NMDA) and transmits excitatory signals. However, in PD, there is an excess release of glutamate in the synaptic cleft which leads to overactivation and sustained activation of NMDA receptors. This overactivation promotes a multifold increase in Ca2+influx within the cell which disturbs the intracellular environment, especially mitochondria. The mitochondria begin to malfunction and release protease which destroys mitochondria. The mitochondrial electron transport chain (ETC) gets disrupted and causes ROS generation. This phenomenon is termed as ‘excitotoxicity’ and it significantly influences the demise of neurons in PD [24, 48,49,50]. Various studies suggest a link between glutamate-mediated excitotoxicity and neuroinflammation in PD. However, there is a controversy that whether neuroinflammation is a detrimental outcome of excitotoxicity or is a cause. Studies suggested that pathological mutations cause changes in the level of glutamate receptors and transporters, implying that excitotoxicity leads to pathology. Excess of glutamate overstimulates glutamatergic receptors of microglial cells which cause neuroinflammation [48, 51]. Contrastingly, some studies suggest that microglial activation releases excessive glutamate stored within them which accelerates neurotoxicity and a strong inflammatory response is observed [52].
S1R activation attenuates glutamate-mediated excitotoxicity in numerous diseases. Selective S1R ligands or non-selective Sigma ligands have shown neuroprotective properties through modulating excitotoxicity. Shimazu et al. tested the effects of one S1R ligand and three sigma ligands on midbrain cell cultures challenged with 100 µM of NMDA. They observed a significant reduction in excitotoxicity and elevation in cell viability using the immunostaining technique. They proposed two mechanisms viz. direct mechanism and indirect mechanism by which these ligands exerted neuroprotective effects. The direct mechanism is related to the binding of sigma receptor ligands to the PCP and NR1/2B site of NMDA receptors and antagonizing them. The indirect mechanism is concerned with the interaction of S1R with the PLc (phospholipase C) pathway (Fig. 3). As the activity of the NMDA receptor is governed by protein kinase C (PKC), indirectly depleting its level may attenuate excitotoxicity. [53, 54]. Decoster et al. reported similar results using rat cortical neurons [55, 56]. Lu et al. has demonstrated that S1R agonist-SKF 10047 ([2S-(2α,6α,11R*]-1,2,3,4,5,6-hexahydro-6,11-dimethyl-3-(2-propenyl)-2,6-methano-3-benzazocin-8-ol) decreases excitotoxicity by modulation of Ca2+ influx (Fig. 3). The SKF 10047 decreases cytosolic Ca2+ levels through N-Type and P/Q-Type calcium channels and thereby decreases glutamate levels. The researchers checked the effects of PKC inhibition on glutamate-mediated excitotoxicity and found that the addition of GF109203X (PKC inhibitor) decreases the glutamate release, verifying the involvement of PKC in mediating excitotoxity [57].
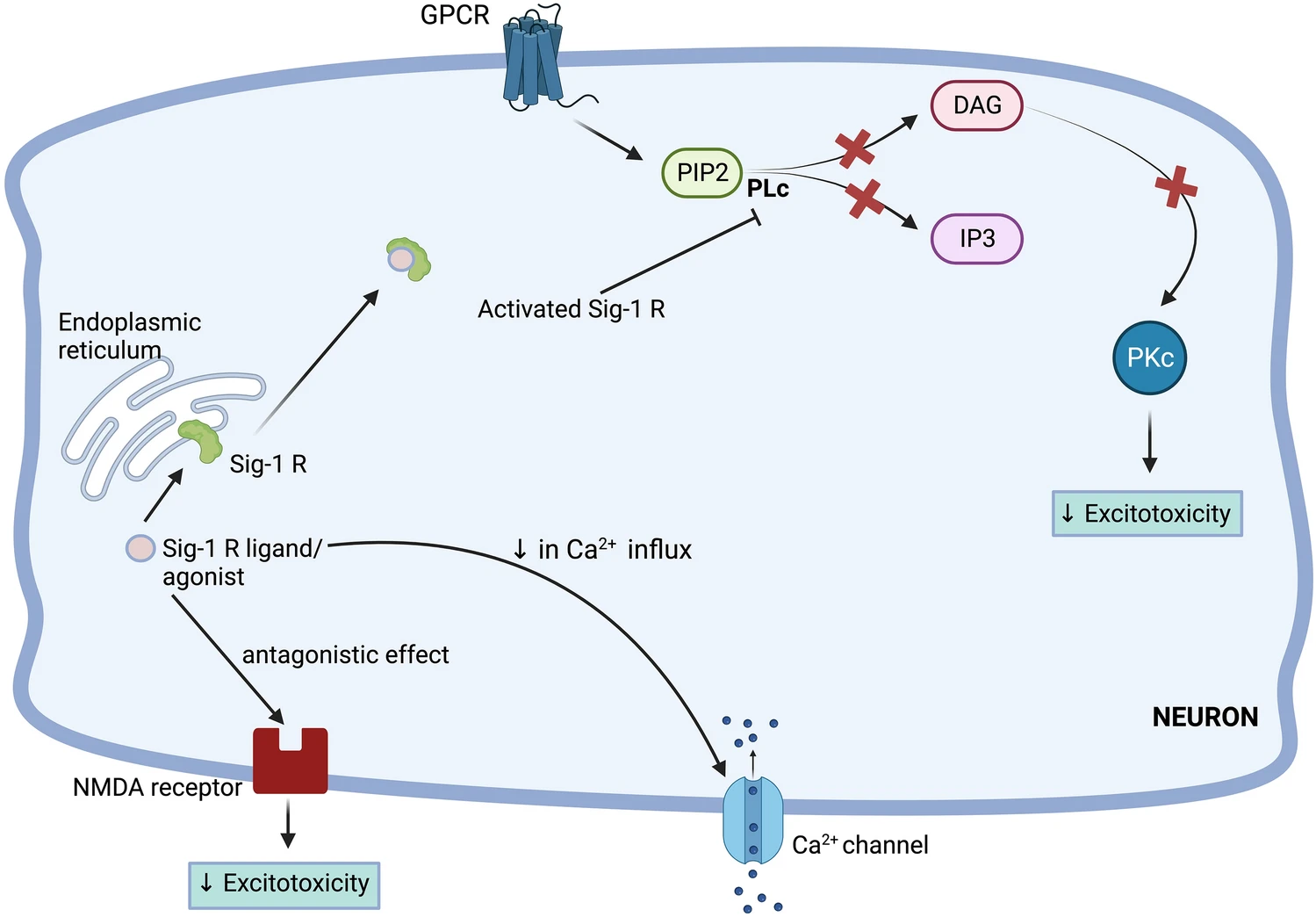
Attenuation of excitotoxicity by sigma 1 receptor
Role of S1Rs in Alleviating Oxidative Stress
ROS are unstable molecules generated by various metabolic reactions that readily react with any component of the cell. They are harmless at physiological concentration and reversely, they are useful under certain cases like a microbial invasion. However, oxidative stress occurs when there is an imbalance between the production and accumulation of ROS and cells fail to combat the detrimental effects of ROS. Oxidative stress is believed to be one of the etiological factors for PD. The ROS reacts with lipids, proteins, and even DNA (deoxyribonucleic acid) which promotes neuronal death. Polyunsaturated fatty acids that are expressed abundantly on the plasma membrane are highly prone to ROS and oxidative stress disrupts the membrane permeability and fluidity. ROS reacts with alpha-synuclein and promotes its aggregation [58]. DNA damage done by ROS disturbs the coding and gene expression. The production of ROS can also be a consequence of a high level of cytosolic dopamine. Excess dopamine can readily be oxidized for the generation of ROS [8, 59,60,61].
S1Rs are reported to protect the cells against oxidative stress. Its activation attenuates ROS accumulation, upregulates endogenous antioxidants, and thereby decreases oxidative stress [24, 25]. Wang et al. reported that S1R activation by (+)-pentazocine attenuated oxidative stress and enhanced cone cell viability. The underlying mechanism proposed by the authors is the modulation of Nrf2 (nuclear factor erythroid 2-related factor). In the absence of oxidative stress, Nrf2 is bounded to KEAP1 (kelch-like ECH-associated protein 1) which enables the degradation of excess Nrf2. During oxidative stress, Nrf2 detaches from KEAP1 and translocates to the nucleus where it binds to ARE (antioxidant response element). ARE is an enhancer sequence found in the promotor region of genes expressing antioxidants [62]. The (+)-pentazocine does not interfere with Nrf2-KEAP1 binding but enhances Nrf2-ARE binding (Fig. 4). (+)-pentazocine increases the expression of Nrf2. The viability of cone cells was lost when (+)-pentazocine was administered to Nrf2 KO mice, verifying the mechanism of S1R ligand [63]. Tuerxun et al. has evaluated the effects of SA4503 (1-(3,4-dimethoxyphenethyl)-4-(3-phenylpropyl) piperazine dihydrochloride)-an S1R agonist on cultured cortical neurons and found that SA4503 inhibited the oxidative stress-induced neuronal death by downregulating MAPK/ERK (mitogen-activated protein kinase/extracellular signal-regulated kinase) pathway (Fig. 4). Cortical neurons when exposed to H2O2-induced oxidative stress lead to overactivation and sustained activation of ERK1/2. Activated ERK 1/2 controls many cellular functions including cell proliferation and cell survival but overactivated ERK1/2 contributes to oxidative stress [64]. Tuerxun et al. has demonstrated that SA4503 decreases the activation of ERK 1/2 and thereby inhibits oxidative stress. Sustained activation of ERK 1/2 leads to glutamate-mediated oxidative toxicity and so the researchers have tested the effect of SA4503 on glutamate receptors. They found that SA4503 downregulates the ionotropic glutamate receptors especially, GluR1 [65]. Tsai et al. has done a complete gene expression analysis of S1R using the Microarray technique. They showed that S1R depletion enhances ROS generation, indicating a direct correlation between S1R and ROS. Their data suggests that S1R deficiency upregulates many antioxidant genes [66].
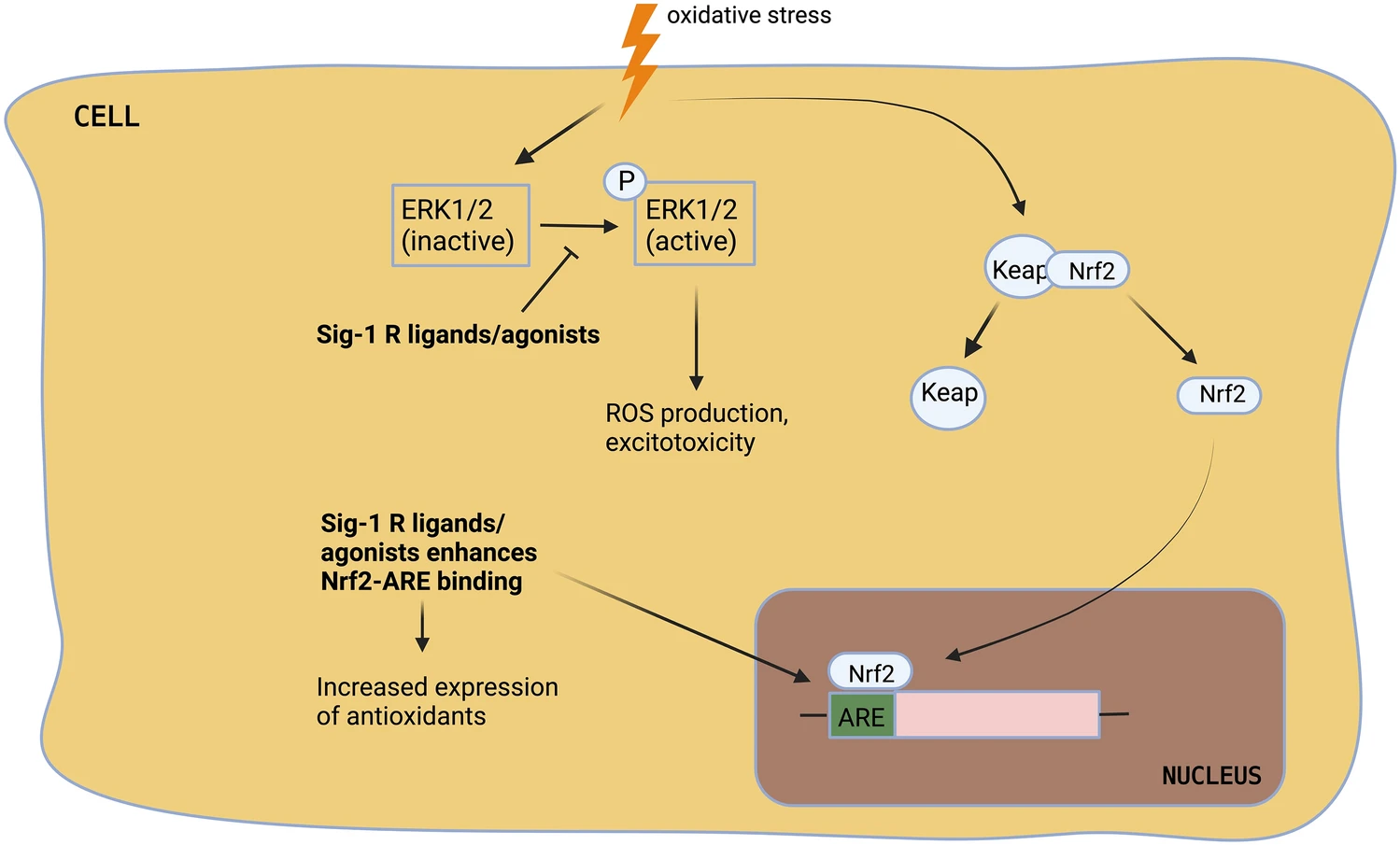
Modulation of ROS by sigma 1 receptor
S1R as a Target for the Treatment of Parkinson’s Disease
The aforementioned sections discussed the role of S1R activation in neuroprotection. Thus, S1R agonists can be potential agents for preventing neurodegeneration. Since the discovery of S1Rs in 1976, their agonists have been tested in-vitro, preclinically and clinically (Tables 1, 2and 3). Wang et al. has used PRE-084—a selective S1R agonist to evaluate its effects on mitophagy in MPTP induced PD model. PRE-084 enhanced neuronal mitophagy and thereby promoted the survival of neurons. The underlying mechanism was identified as S1R mediated mitophagy through PINK1/parkin pathway. The mitophagy is governed by PINK1 and activated S1R stabilizes PINK1 leading to the enhancement of mitophagy [67]. Francardo et al. used the same S1R agonist i.e. PRE-084 to study its neuroprotective properties. They used male C57BL/6 mice for the experimental study and PD was induced by stereotaxically injecting 6-hydroxydopamine (6-OHDA) into basal ganglia. Chronic treatment (5 weeks) with PRE-084 (0.3 or 1 mg/kg; s.c.) improved motor impairment in mice, evaluated by cylinder, stepping, and spontaneous rotational activity tests. There was a significant enhancement in TH+ cells in the substantia nigra region. The microglial activation was reduced and dopamine levels were elevated in the treatment group (Table 1). Studies on the impact of S1R KO on the activity of PRE-084 showed that the activity was lost in S1R KO mice [41]. In 2019, Francardo et al. again conducted the same experimental study with a different S1R agonist pridopidine [42]. Pridopidine is reported to improve motor abnormalities in an animal model of Huntington’s disease [68]. The pridopidine at doses of 0.3 or 1 mg/kg s.c. showed similar neuroprotective and neurorestorative effects as observed with PRE-084. Pridopidine enhanced the neurotrophic factors like GDNF (glial cell neurotrophic factor), BDNF (brain-derived neurotrophic factor), and ERK1/2. Similarly, the activity of pridopidine was lost in S1R KO mice [42].
| Sr. No | S1R agonist | Animal model | Dose (in mice) | Outcome/Remarks | Refs |
|---|---|---|---|---|---|
| 1) | PRE-084 | 6-OHDA, MPTP | 0.3 or 1 mg/kg | • Restoration of mitophagy and clearance of damaged mitochondria• Motor recovery in C57BL/6 mice• Enhancement in TH+ cells and dopamine levels• Reduction in microglial activation | [28, 65] |
| 2) | Pridopidine | 6-OHDA | 0.3 or 1 mg/kg | • Enhancement in cell viability• Enhancement in neurotrophic factors (GDNF, BDNF and ERK1/2)• Enhancement in TH+ cells and dopamine levels | [39] |
| 3) | Afobazole | 6-OHDA | 2.5 mg/kg | • Improvement in falling latency in rotarod test• Increase in striatal dopamine content | [67, 68] |
Preclinical studies of some S1R agonists in animal models of Parkinson’s disease
| Sr. No | S1R agonist | Major outcome | Refs |
|---|---|---|---|
| Preclinical studies | |||
| 1) | 2-(4-Morpholino) ethyl-1-phenylcyclohexane-1-carboxylate(PRE-084) | Modulation of neuroinflammation• The S1R agonistic activity was studied using embolic stroke model in rats• PRE-084 (5 mg/kg; i.p.) improved motor performance of rats as assessed by beam walk rats• The levels of pro-inflammatory cytokines (TNF-α, IL-1α and IL-1β) were reduced and anti-inflammatory cytokines (IL-4 and IL-6) were elevated | [45] |
| 2) | Dextromethorphan | Modulation of excitotoxicity• The ibotenate induced brain injury model was used to study S1R agonistic activity in developing mice brains• Dextromethorphan reduced the neuron damage produced by ibotenate• The NMDA mediated excitotoxicity was less in dextromethorphan treated groups• The mechanism suggested by researchers is antagonism of NMDA receptor by this S1R agonist | [71] |
| 3) | tetrahydro-N,N-dimethyl-2,2-diphenyl-3-furanmethanamine hydrochloride (ANAVEX2-73) | Modulation of oxidative stress• The S1R agonistic activity was studied in Swiss OF-1 mice• The ANAVEX2-73 attenuated ROS levels generated by pathogenic amyloid β• The complex I and IV of ETC was increased suggesting enhanced mitochondrial respiration• The suggested mechanism could be modulation of ERK pathway or blockade of Ca2 +—activated K + (SK) channels | [72] |
| Clinical studies | |||
| 1) | SA4503(Cutamesine) | • A Phase IIa, randomized, double-blinded study was conducted for acute ischemic stroke• 1 mg/day and 3 mg/day of SA4503 administered orally was safe and well tolerated• No drug related adverse events were reported• However therapeutically no significant changes were observed/reported | [73] |
Preclinical and clinical studies of S1R agonists offering neuroprotection through modulation of misfolded protein-induced ER stress, neuroinflammation, excitotoxicity, and oxidative stress
| Sr. No | S1R agonist | In-vitro studies | Refs |
|---|---|---|---|
| 1) | Pridopidine | Modulation of misfolded protein-induced ER stress• The S1R agonistic activity was studied in murine striatal cell line• Pridopidine reduced the levels of UPR sensor proteins i.e. IRE1, PERK and ATF6 which all gets elevated during ER stress | [74] |
| 2) | Afobazole | Modulation of neuroinflammation• The S1R agonistic activity was studied in rat primary microglial cultures• Afobazole inhibited the activation and migration of microglial cells as checked by migration assay• The mechanism explained by researchers is antagonism of P2X and P2Y receptors expressed on microglial cells• The antagonism inhibited microglial activation and response | [69] |
| 3) | (+)-N-allylnormetazocine ((+)-SKF 10047) | Modulation of excitotoxicity• The S1R agonistic activity was studied on midbrain cell cultures• (+)-SKF 10047 improved the neuronal viability and decreased NMDA-mediated excitotoxicity• (+)-SKF 10047 directly binds to the NMDA receptor site and antagonizes them | [53] |
| 4) | SA4503(Cutamesine) | Modulation of oxidative stress• The S1R agonistic activity was studied in cultured cortical neurons• SA4503 improved the neurons survival as compared to control• The underlying mechanism is inhibition of MAPK/ERK pathway which is involved in generating ROS | [65] |
In-vitro studies of some S1R agonists offering neuroprotection through modulation of misfolded protein-induced ER stress, neuroinflammation, excitotoxicity, and oxidative stress
Voronin et al. has demonstrated the neuroprotective effects of afobazole- an anxiolytic with S1R agonistic property. Parkinsonism was induced in ICR mice using 6-OHDA by stereotaxy. The mice treated with 2.5 mg/kg i.p. of afobazole for 2 weeks (either beginning from the 1st day of 6-OHDA injection or the 14th day of 6-OHDA injection) enhanced TH+ cells count and the latency to fall in the rotarod test. Using HPLC-ED (high-performance liquid chromatography-electrochemical detector), striatal dopamine content was measured and was found to be increased in the treatment group (Table 1). The activity of afobazole was lost when BD-1047 (Sig-1 R antagonist) was administered before afobazole administration [69, 70]. Due to these properties, the afobazole was selected for clinical trials. In 2019, Phase II trials commenced investigating its safety and effectiveness for the treatment of levodopa-induced dyskinesia in patients with PD (Clinical trial ID: NCT03922711). A total of 23 participants were recruited and the trial was double-blinded and randomized. However, due to coronavirus disease 2019 (COVID-19) pandemic, the trial was terminated and no significant data was collected.
Conclusion
In PD, neurodegeneration occurs because of numerous pathophysiological factors like pathogenic alpha-synuclein aggregation, neuroinflammation, excitotoxicity, ROS, etc. Activation of S1Rs by agonists provides neuroprotection via different mechanisms. S1R KO mice have shown increased pathogenic alpha-synuclein aggregation and S1R agonists have shown increased degradation of alpha-synuclein. This confirms the role of S1R in alpha-synuclein aggregation. In addition, S1R activation decreases microglial activation and inflammatory response. S1R agonists antagonize NMDA receptors thus inhibiting glutamate-mediated excitotoxicity. Further, S1R agonists modulate gene expression of endogenous antioxidants. All these properties make S1R an ideal therapeutic target for PD. S1R agonists have shown promising results in animal models of PD. Although the results of preclinical studies look promising, detailed clinical studies are required. A thorough investigation will aid in dragging the molecules from the bench to the bedside.