
Sigma-1 receptor modulation fine-tunes KV1.5 channels and impacts pulmonary vascular function

By Alba Vera-Zambrano, Maria Baena-Nuevo, Susanne Rinné, Marta Villegas-Esguevillas, Bianca Barreira, Gokcen Telli, Angela de Benito-Bueno, José Antonio Blázquez, Belén Climent, Francisco Pérez-Vizcaino, Carmen Valenzuela, Niels Decher, Teresa Gonzalez, and Angel Cogolludo
Excerpt from the article published in Pharmacological Research, Volume 189, March 2023, 106684,
ISSN 1043-6618, DOI: https://doi.org/10.1016/j.phrs.2023.106684.
Editor’s Highlights
- KV1.5 channels are responsible for setting or regulating the resting membrane potential and thereby, for controlling pulmonary vascular tone, cell apoptosis, proliferation, and survival of pulmonary artery smooth muscle cells (PASMC).
- Sigma-1 receptor (S1R) associates to and regulates several K+ channels, including hERG, KV1.2, KV1.3, KV1.4 and KV1.5.
- Pulmonary arterial hypertension and atrial fibrillation are severe cardiovascular diseases associated with electrical remodelling including KV1.5 channel dysfunction.
- Chaperone S1R functionally interacts with the KV1.5 channel and acts as a critical regulator of its expression and activity. In the arterial pulmonary vessels, S1R agonists are able to increase KV1.5 currents and reduce contraction and proliferation of PASMC.
- S1R is a novel pharmacological target to rescue the attenuated KV1.5 channel function in cardiovascular diseases associated with channel malfunction.
Abstract
KV1.5 channels are key players in the regulation of vascular tone and atrial excitability and their impairment is associated with cardiovascular diseases including pulmonary arterial hypertension (PAH) and atrial fibrillation (AF). Unfortunately, pharmacological strategies to improve KV1.5 channel function are missing. Herein, we aimed to study whether the chaperone sigma-1 receptor (S1R) is able to regulate these channels and represent a new strategy to enhance their function. By using different electrophysiological and molecular techniques in X. laevis oocytes and HEK293 cells, we demonstrate that S1R physically interacts with KV1.5 channels and regulate their expression and function. S1R induced a bimodal regulation of KV1.5 channel expression/activity, increasing it at low concentrations and decreasing it at high concentrations. Of note, S1R agonists (PRE084 and SKF10047) increased, whereas the S1R antagonist BD1047 decreased, KV1.5 expression and activity. Moreover, PRE084 markedly increased KV1.5 currents in pulmonary artery smooth muscle cells and attenuated vasoconstriction and proliferation in pulmonary arteries. We also show that both KV1.5 channels and S1R, at mRNA and protein levels, are clearly downregulated in samples from PAH and AF patients. Moreover, the expression of both genes showed a positive correlation. Finally, the ability of PRE084 to increase KV1.5 function was preserved under sustained hypoxic conditions, as an in vitro PAH model. Our study provides insight into the key role of S1R in modulating the expression and activity of KV1.5 channels and highlights the potential role of this chaperone as a novel pharmacological target for pathological conditions associated with KV1.5 channel dysfunction.
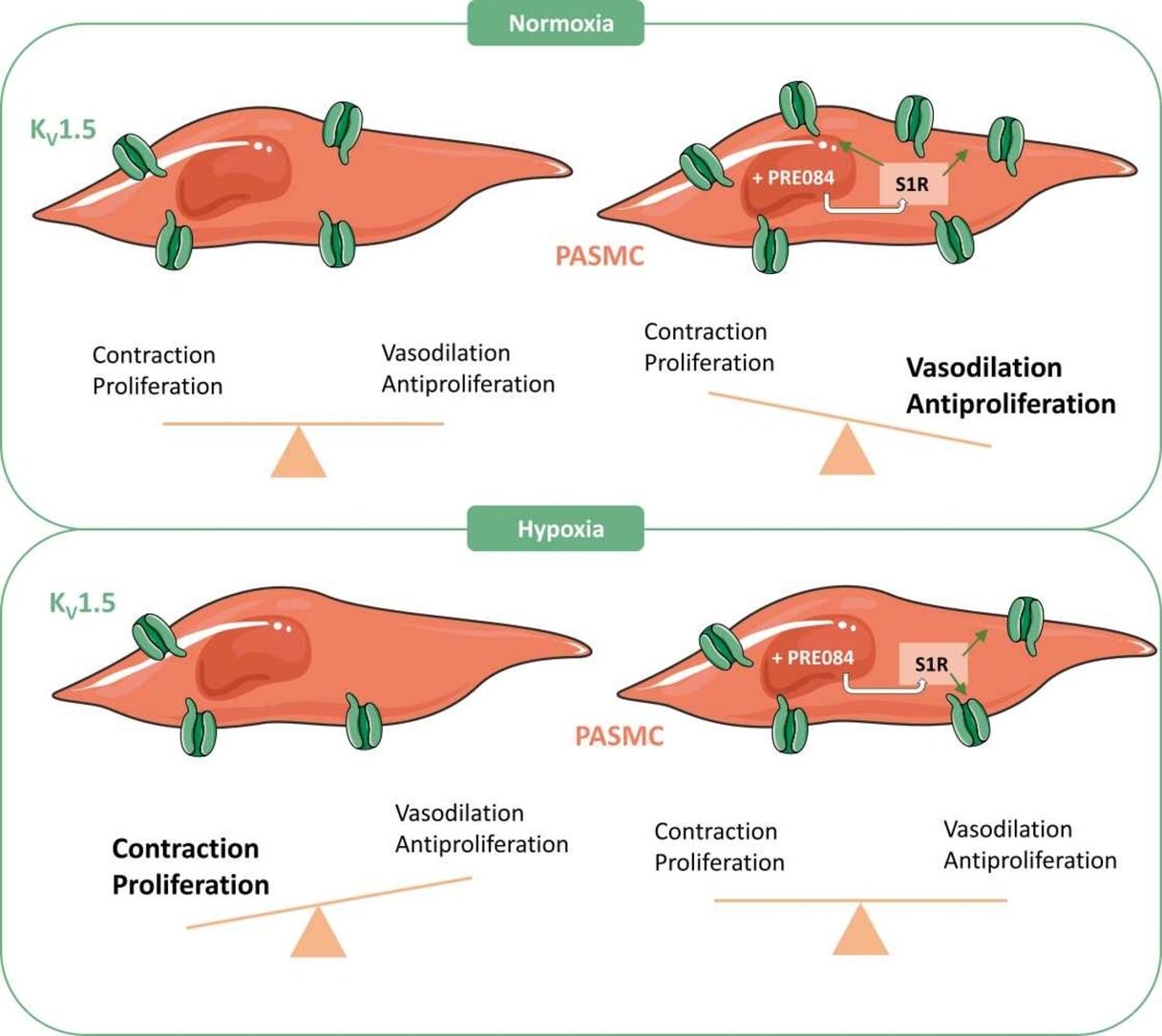
1. Introduction
KV1.5 channels play a key role in the pulmonary circulation. They are responsible for setting or regulating the resting membrane potential and thereby, for controlling pulmonary vascular tone, cell apoptosis, proliferation, and survival of pulmonary artery smooth muscle cells (PASMC) [1], [2], [3], [4], [5]. Moreover, KV1.5 dysfunction is a common feature in pulmonary arterial hypertension (PAH) [4], [5], [6]. In the cardiovascular system, KV1.5 channels also participate in the cardiac atrial repolarization of the action potentials [7] and their dysregulation is a common feature in atrial fibrillation (AF). Therefore, KV1.5 channels are considered potential therapeutic targets in these pathological conditions, but unfortunately, pharmacological strategies to rescue the reduced expression and activity of KV1.5 channels are still lacking.
Sigma-1 receptor (S1R) is a 25-KDa chaperone which plays a central role in protein folding and quality control [8]. In a resting state, it resides in the mitochondrion-associated membrane (MAM) where it forms a complex with a chaperone called immunoglobulin protein (BiP) [8]. When activated, S1R interacts with different client proteins in the ER or other organelles [9] and, under chronic stress, it can translocate to the plasma membrane to regulate the activity of different plasma membrane proteins such as ion channels [10], [11], [12], [13].
S1R can interact with itself to form oligomers and their distribution can be modified by different ligands. In this regard, agonists (e.g., SKF10047, PRE084) bias the equilibrium to lower molecular weight states while antagonists (e.g., BD1047, haloperidol, and sphingosine) may shift the equilibrium to higher molecular weight states [14], [15]. Moreover, those ligands that induce hyperlocomotion or other physiological responses through binding to S1R were considered as agonists, and those blocking or blunting these responses were considered as antagonists [9], [16].
Due to its role in the human physiology, the function of S1R has been studied in different pathologies and, nowadays, it is considered as a potential therapeutic target for several disorders including neurodegenerative pathologies such as Alzheimer’s and Parkinson’s diseases [17], stroke [18] and cancer [19]. During the last decade, several groups have suggested that S1R represents a novel molecular target in cardiovascular diseases [20]. S1Rs are expressed in cardiomyocytes [21], [22], and their ligands have shown to alter contractility, calcium flow and rhythmic activity [23], [24]. Previous studies suggest that S1R associates to and regulates several K+ channels, including hERG, KV1.2, KV1.3, KV1.4 and KV1.5 [10], [12], [13], [25], [26]. Few in vitro studies in heterologous systems suggest that KV1.5 channels can be targeted by S1R ligands, but it is still unknown whether S1R regulates K+ channels, in particular, KV1.5 in the cardiovascular system and whether this interaction can be modulated pharmacologically.
Therefore, the aim of this study was to analyse the potential modulation of KV1.5 channels by S1R and its possible physiological, pathophysiological and pharmacological significance. Our data identify S1R as a potential novel pharmacological target in pathological conditions associated with KV1.5 impairment.
…
3. Results
3.1. Bimodal modulation of KV1.5 channels by S1R
To start the characterization of the potential interaction between S1R and the KV1.5 channel we first studied the potassium currents recorded by TEVC in X. laevis oocytes transfected with a single KV1.5 and different S1R cRNA concentrations. Fig. 1A shows representative overlapping current traces at different S1R concentrations. Interestingly, we found that low S1R concentrations (KV1.5:S1R ratio 1:0.2) increased, while higher concentrations (1:2) decreased KV1.5 current amplitude (Fig. 1B).
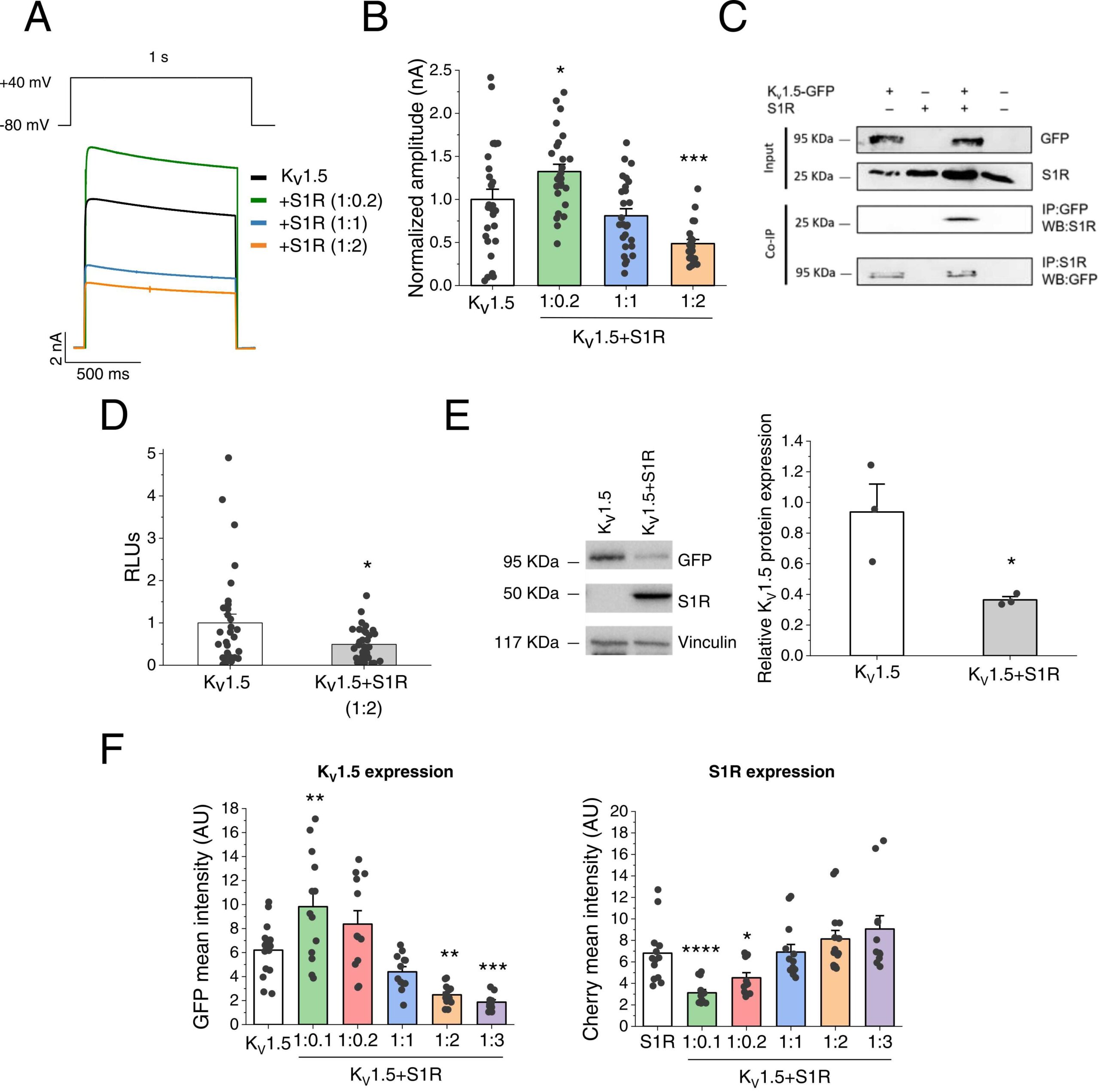
Modulation of KV1.5 by S1R.
(A) Representative current traces of KV1.5 and KV1.5 +S1R at different KV1.5:S1R ratios (1:0.2, 1:1, 1:2) recorded in X.laevis oocytes. (B)Maximum current amplitude of KV1.5 and KV1.5 +S1R at different ratios, 48 h after injection. Maximum current at + 40 mV (nA) in all conditions was normalized to KV1.5 amplitude (n = 22–28 oocytes, from N = 3 X.laevis). (C) Protein extracts were prepared from HeLa cells (input). The Co-IP was performed by immunoprecipitating with an anti-GFP antibody and probing with an anti-S1R antibody (n = 2). To confirm the interaction, a reverse Co-IP experiment immunoprecipitating with an anti-S1R antibody and probing with an anti-GFP antibody was performed (n = 1). (D) Protein expression of KV1.5HA in the plasma membrane of X. laevis oocytes injected with KV1.5HA or KV1.5HA+S1R (1:2 ratio). RLUs: relative light units (n = 32 oocytes, from N = 2 X.laevis). (E) Relative protein expression of KV1.5 (GFP+) and S1R in KV1.5 and KV1.5 +S1R conditions, analysed by western blot in HEK293 cells and normalized by vinculin expression (n = 3). (F) Mean intensity of the GFP and cherry signals (arbitrary units: AU) by flow cytometry in HEK293 cells at different KV1.5:S1R ratios (n = 12–15). * : P < 0.05, * *: P < 0.01, * ** : P < 0.001 and * ** *: P < 0.0001 vs. KV1.5. In panel B and F right vs. S1R. In panels B and D, data were analysed by a 1-way ANOVA followed by a post-hoc Dunnett test, in D and E, by a two-tailed unpaired student t-test.
Next, we searched for potential evidence of physical protein-protein interaction between KV1.5 and S1R by Co-IP studies. To this end, HeLa cells co-transfected with KV1.5 and S1R were immunoprecipitated with the anti-GFP antibody and probed with the anti-S1R antibody, consistent with the Co-IP of the two proteins. This protein-protein interaction was confirmed by a reverse immunoprecipitation with the anti-S1R antibody and by probing with the anti-GFP antibody (Fig. 1C).
Since changes in current amplitude may be essentially due to alterations in gating and/or expression of KV1.5 channels in the plasma membrane, we studied this second possibility in X. laevis oocytes injected with KV1.5, now harbouring an extracellular HA-epitope, alone or with S1R (ratio KV1.5-S1R 1:2). Then, the surface expression was probed with ELISA assays, exclusively detecting channels at the plasma membrane. Our data revealed that the decreased current amplitude at that ratio was linked to a decreased surface expression of KV1.5 channels (Fig. 1D). A similar downregulation of total KV1.5 channel expression was observed in HEK293 cells transfected with a 1:2 KV1.5:S1R ratio by Western blot (Fig. 1E) and in HeLa cells by fluorescence microscopy (see Supplementary material online, Fig. S1). Interestingly, S1R did not affect any electrophysiological feature apart from the referred decrease in current amplitude (see Supplementary material online, Table S1) suggesting that this chaperone mainly affects the expression rather than the gating of KV1.5 channels. To analyse whether S1R also exerts a bimodal regulation of KV1.5 channel expression, KV1.5-GFP and S1R-cherry plasmids were transfected at different ratios in HEK293 cells, and the mean fluorescence signal of both proteins was determined by flow cytometry. Our data confirmed that overexpression with low S1R levels was associated with increased total KV1.5 channel expression while higher S1R concentrations led to downregulation of KV1.5 channels (Fig. 1F).
Finally, to elucidate whether the modulation by S1R was specific for KV1.5 channels, we also analysed its effects on KV2.1 and Kir2.1 channels in X. laevis oocytes. We found that current amplitude recorded at + 40 mV was similar for KV2.1 (2.48 ± 0.30 nA, n = 14) and KV2.1 +S1R (2.41 ± 0.26 nA, n = 14; P > 0.05) (see Supplementary material online, Fig. S2A, C). Similarly, Kir2.1 current amplitude recorded at −140 mV (1.40 ± 0.13 nA, n = 14) was not significantly affected by S1R (1.22 ± 0.11 nA, n = 17; P > 0.05) (see Supplementary material online, Fig. S2B-C).
3.2. Incubation with S1R ligands modulate KV1.5 channels
Next, we studied whether S1R ligands could modulate KV1.5 channel expression and activity. We found that acute exposure (∼ 10 min) to the agonist PRE084 led to a reduction (< 20 %) of the potassium current amplitude that was similar in HEK293 cells transfected with KV1.5 or KV1.5 +S1R (see Supplementary material online, Fig. S3A, B). Notably, a similar inhibitory effect was observed after acute challenge to the S1R antagonist BD1047, which was also unaltered by S1R expression (see Supplementary material online, Fig. S3C, D).
Since our data suggested that S1R affects mainly the expression rather than the gating of KV1.5 channels we then focused on the sustained (24 h) effects of PRE084. KV1.5 channel activity assessed by whole-cell patch-clamp markedly augmented following the incubation with PRE084 in HEK293 cells transfected with KV1.5 +S1R (2.55 ± 0.26 nA, n = 36 vs. 3.44 ± 0.28 nA, n = 33 at + 40 mV, in the absence and in the presence of PRE084, respectively; P < 0.05) (Fig. 2A, B). Of note, this enhancing effect of PRE084 was not observed in cells only transfected with the channel (i.e. in the absence of S1R; see Supplementary material online, Fig. S4A-B).
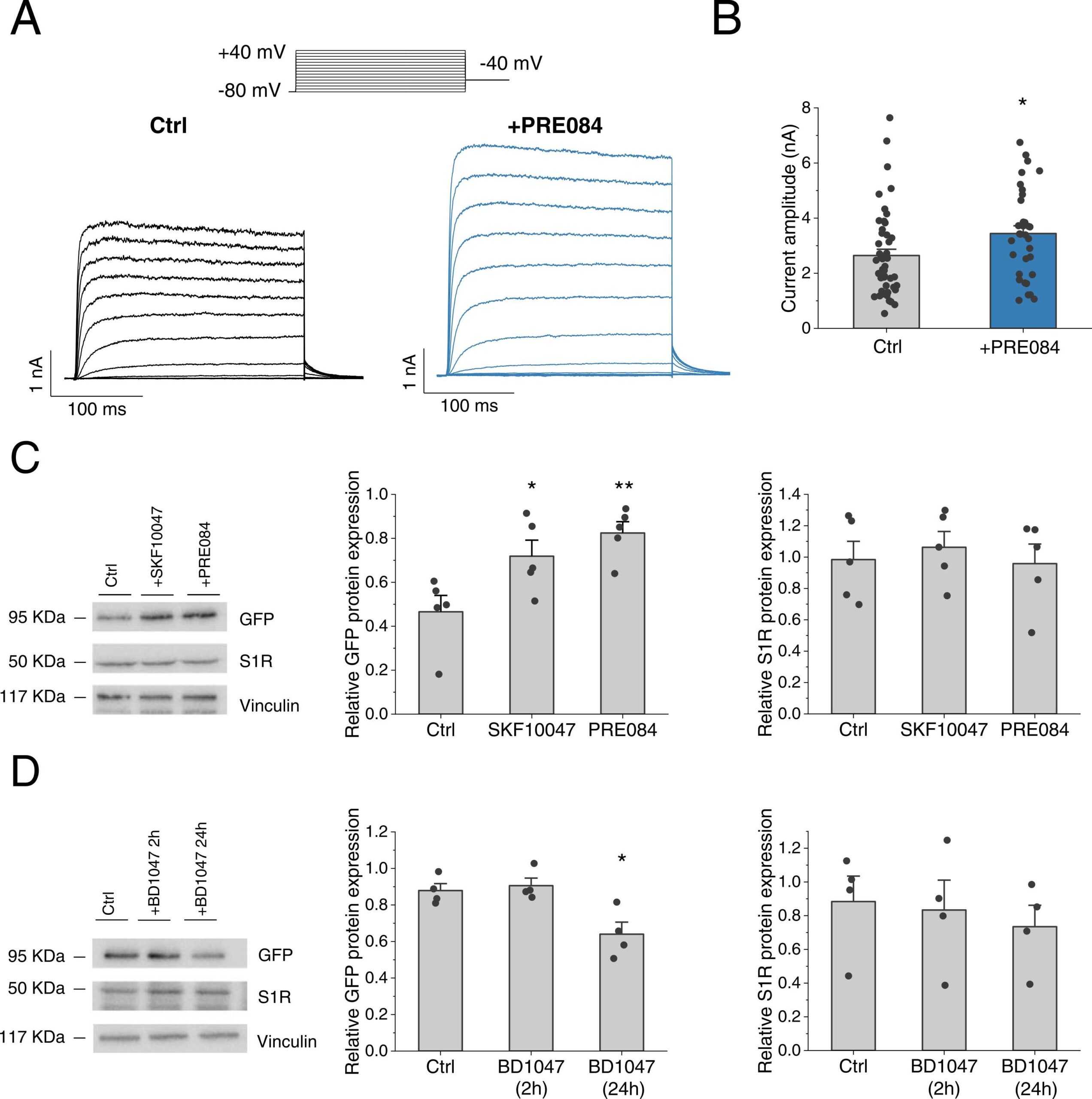
Modulation of KV1.5 activity and expression by S1R ligands in HEK293 cells.
(A)Representative I-V recordings and (B) average data of the maximum KV1.5 current amplitude at + 40 mV was analysed in cells transfected with KV1.5 and S1R after incubation for 24 h with (n = 33) or without PRE084 (10 µM) (n = 47). (C) Cells transfected with KV1.5 +S1R were incubated for 24 h with PRE084 (20 µM) or SKF10047 (20 µM) (n = 5). (D) Cells transfected with KV1.5 +S1R were incubated with BD1047 (10 µM) for 2 or 24 h (n = 4). In B and C panels, relative KV1.5 (left) and S1R protein expression (right) by western blot was probed with an anti-GFP or anti-S1R antibody and normalized by vinculin expression. * : P < 0.05, * *: P < 0.01 vs. control. In panels B, data was analysed by a two-tailed unpaired student t-test. In C and D, by a 1-way ANOVA followed by a post-hoc Dunnett test.
Protein expression analysis revealed that 24 h incubation with the S1R agonists PRE084 or SKF10047 led to a marked increase in KV1.5 expression (Fig. 2C left). Conversely, chronic (24 h) but not acute (2 h) incubation with the S1R antagonist BD1047 reduced KV1.5 expression (Fig. 2D left). Neither of these modulators significantly affected the S1R expression (Fig. 2C–D, right panels). Of note, the effect of PRE084 on KV1.5 expression was not observed in cells not transfected with S1R (see Supplementary material online,Fig. S4C) strongly suggesting that PRE084 effect was though S1R.
3.3. The S1R agonist PRE084 increases KV1.5 current and decreases contraction in pulmonary arteries
Since KV1.5 channels play an important role in the pulmonary circulation we aimed to study the impact of S1R modulation on these channels in rat isolated PASMC. Whole-cell patch-clamp recordings from PASMC revealed that the incubation during 24 h with PRE084 dramatically increased the total K+ current (Fig. 3A). To analyse whether the KV1.5 current, which represents the main component of the total KV current in the pulmonary vasculature, was affected by PRE084, cells were perfused with DPO-1 (1 µM), a selective KV1.5 channel inhibitor. DPO-1-sensitive currents, which reflect the KV1.5 channel component, were markedly higher in PASMC following PRE084 (20 µM) treatment (54.15 ± 10.03 pA/pF at +40 mV, n = 9) in comparison with those observed in PASMC treated with vehicle (27.61 ± 2.91 pA/pF, n = 8; P < 0.05) (Fig. 3B left). A further increase in KV1.5 currents was observed when PA were incubated with a higher concentration of PRE084 (50 µM), yielding values of 128.29 ± 18.97 pA/pF at + 40 mV (n = 6; P < 0.01 vs Control; see Supplementary material online, Fig. S5). On the other hand, PRE084 (20 µM) did not significantly affect the DPO-1-insensitive current confirming that the increase in K+ current was mainly due to KV1.5 channels (Fig. 3B right). PRE084 did not modify the voltage-dependence activation (Fig. 3C) but induced a lower degree of inactivation in the total current than that observed in control PASMC (3.53 ± 1.74 % and 17.08 ± 5.82 %, respectively, n = 10; P < 0.05) (Fig. 3D).
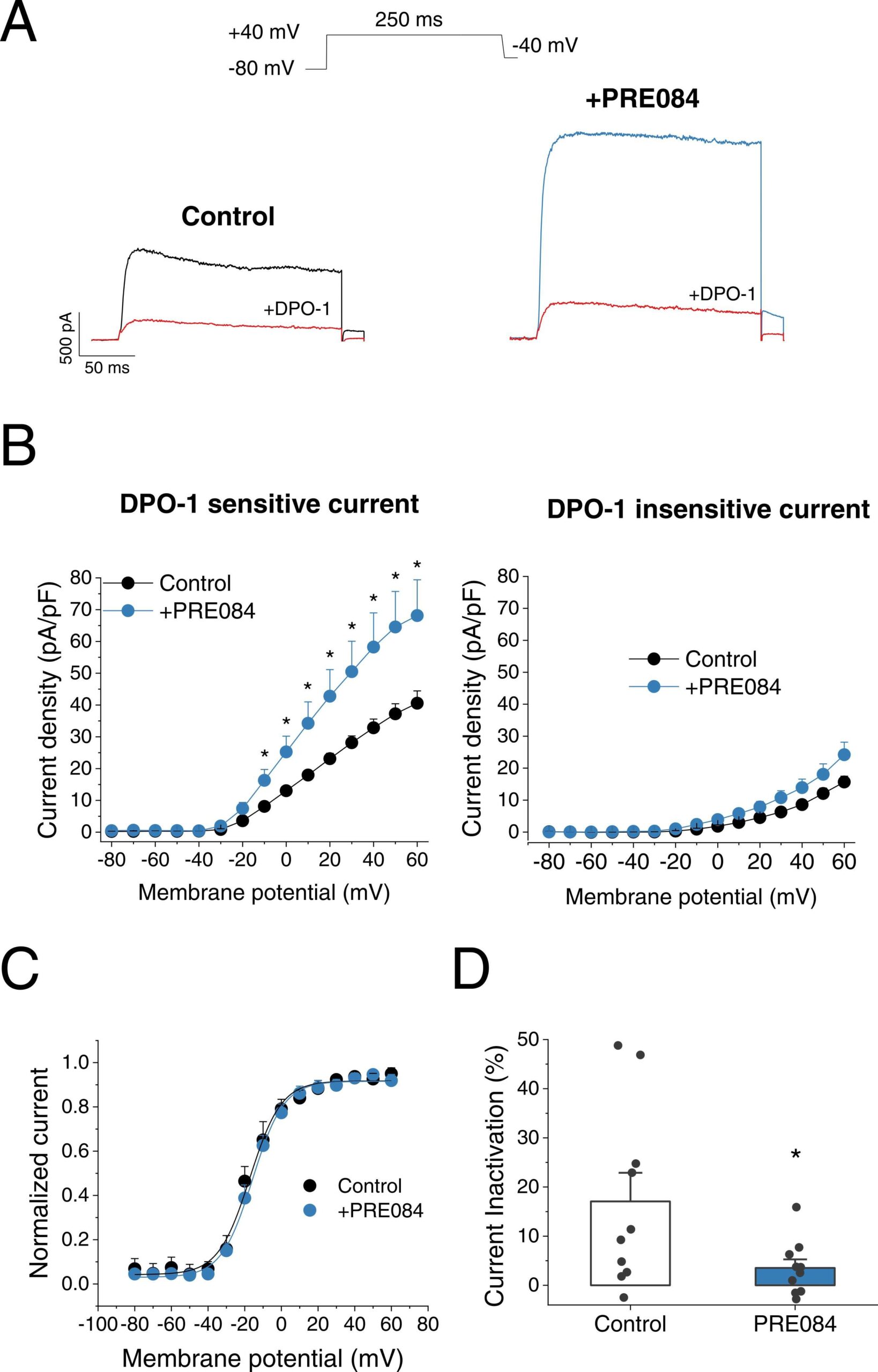
Effect of PRE084 on K+ currents in PASMC.
(A) Representative current traces of K+current at + 40 mV in control condition (left, black) and incubated with PRE084 (20 µM) for 24 h (right, blue). The current was register in the absence and in the presence of DPO-1 (red). (B) I-V relationships obtained after plotting the maximum current amplitude, normalized by cell capacitance, at each membrane potential (n = 8–9 cells from N = 6 rats). The DPO-1 sensitive current is showed on the left and the DPO-1 insensitive current on the right. (C) Normalized activation curves in control condition and after incubation with PRE084 (20 µM) (n = 9 from N = 6 rats per group). (D) Degree of inactivation of the current in control condition and after incubation with PRE084 (20 µM) (n = 10 from N = 6 rats per group). * : P < 0.05 vs. control. Data were analysed by a two-tailed unpaired student t-test.
As mentioned in the introduction, KV1.5 channel activity/expression critically regulates PA contractility. Therefore, the effects of 24 h incubation with PRE084 (20 µM) on the contractility of these vessels were studied. The vasoconstrictor response to KCl (80 mM), which is regarded as an index of the contractile capacity of the vessel, were unaffected by incubation with 20 µM PRE084 (1.84 ± 0.22 mN/mm2, n = 12 vs. 1.64 ± 0.23 mN/mm2, n = 9). On the other hand, PRE084 decreased PA contraction to Phe (Fig. 4A–B). In particular, PRE084 reduced the maximal contraction (Emax), but not the potency, of Phe in a concentration-dependent manner (see Supplementary material online, Table S2). Moreover, the attenuated Phe contraction by PRE084 (20 µM) was not observed in the presence of DPO-1 (Fig. 4C–D), strongly suggesting that the attenuation of pulmonary vasoconstriction induced by PRE084 was mainly due to an increase in KV1.5 channel activity. In addition, to test whether PRE084 influences cell proliferation or apoptosis, human PASMC were incubated with increasing concentrations of PRE084 (1, 20 and 50 µM) for 24 h. Our results showed that PRE084 decreased PASMC proliferation in a concentration-dependent manner (Fig. 4E). On the other hand, this drug did not significantly affect cell viability as assessed by flow cytometry analysis of annexin-V and propidium iodide staining of human PASMC (see Supplementary material online, Fig. S6).
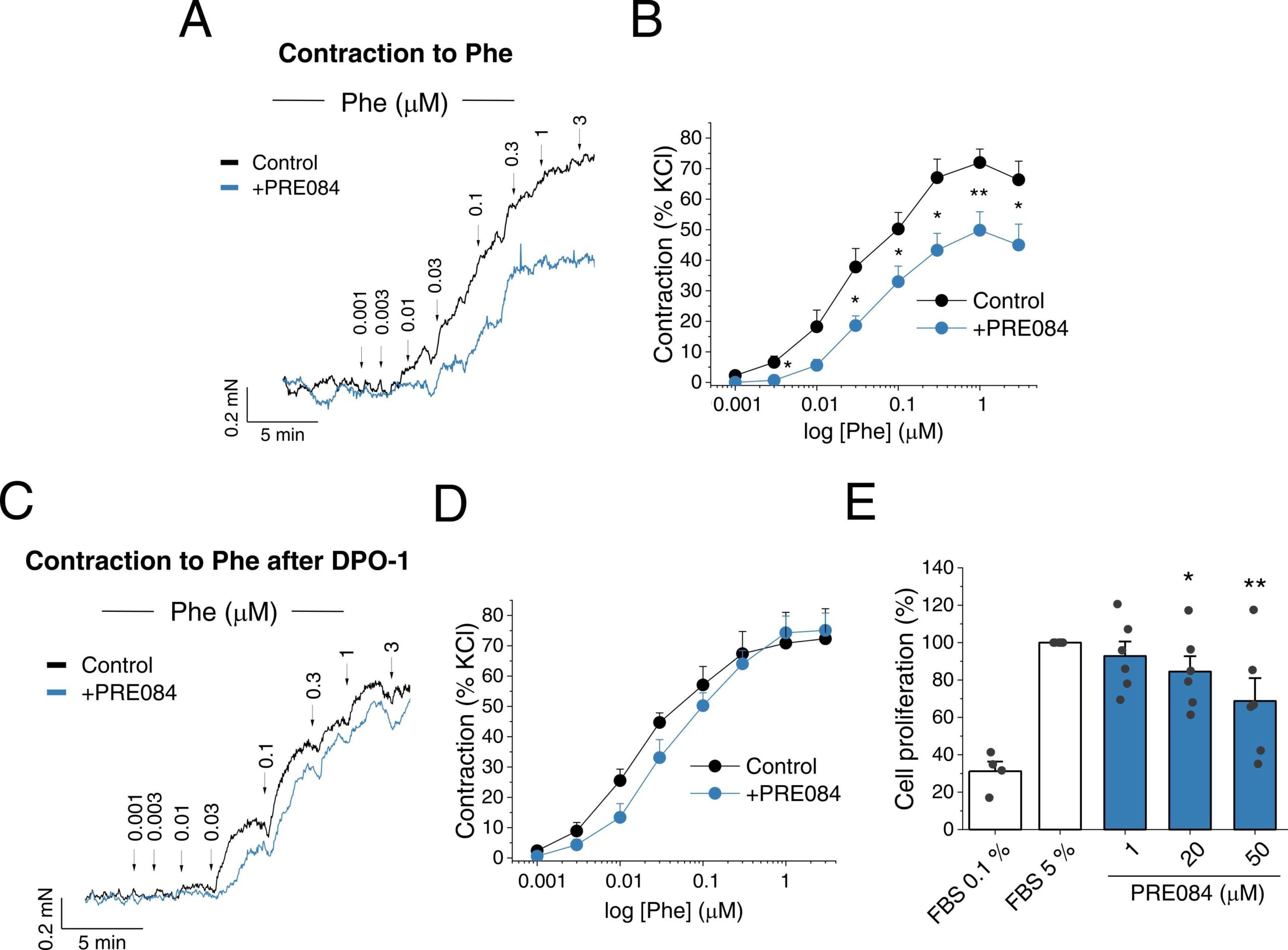
PRE084 decreases vasoconstriction and proliferation in PA.
(A) Representative recording and (B) average data of the concentration-response curves to phenylephrine (Phe, 0.001 – 3 µM) in rat PA incubated for 24 h in the absence (control) or in the presence of PRE084 (20 µM) (n = 12 PA from N = 8 rats per group). (C) Representative recording and (D) average data of the concentration-response curves to Phe after 15 min treatment with DPO-1 in PA incubated for 24 h in the absence (control) or in the presence of PRE084 (20 µM) (n = 9 PA from N = 6 rats per group). (E) Concentration-dependent inhibition of human PASMC proliferation induced by PRE084 (1, 20 and 50 µM) (n = 6 from N = 5 patients). Data were normalized to the control condition (FBS 5 %). FBS 0.1 % was used as a negative control of proliferation (n = 4). *: P < 0.05, **: P < 0.01 vs. control. Data were analysed by a two-tailed unpaired student t-test (panel B) or 1-way ANOVA followed by a post-hoc Dunnett test (panel E).
3.4. The expression of KV1.5 and S1R is decreased in PAH and AF
Next, we aimed to determine whether the expression of S1R was impaired in cardiovascular diseases associated with altered KV1.5 channel expression such as PAH and AF. Regarding PAH, first we performed a transcriptomic analysis which revealed that, both KCNA5 and SIGMA1R mRNA levels were clearly downregulated in samples from PAH patients (Fig. 5A). Moreover, the expression of both genes showed a positive correlation, with a P = 0.038 (Fig. 5B). When the data was stratified according to PAH types, a similar downregulation was observed in all types of PAH studied, although some data did not reach statistical significance, probably due to the low number of samples (Fig. 5C). In line with these data, we found that KV1.5 and S1R protein expressions were also downregulated in lung samples from PAH patients compared to controls (Fig. 5D). While both proteins exhibited a subtle positive correlation trend this did not reach statistical significance (Fig. 5E).
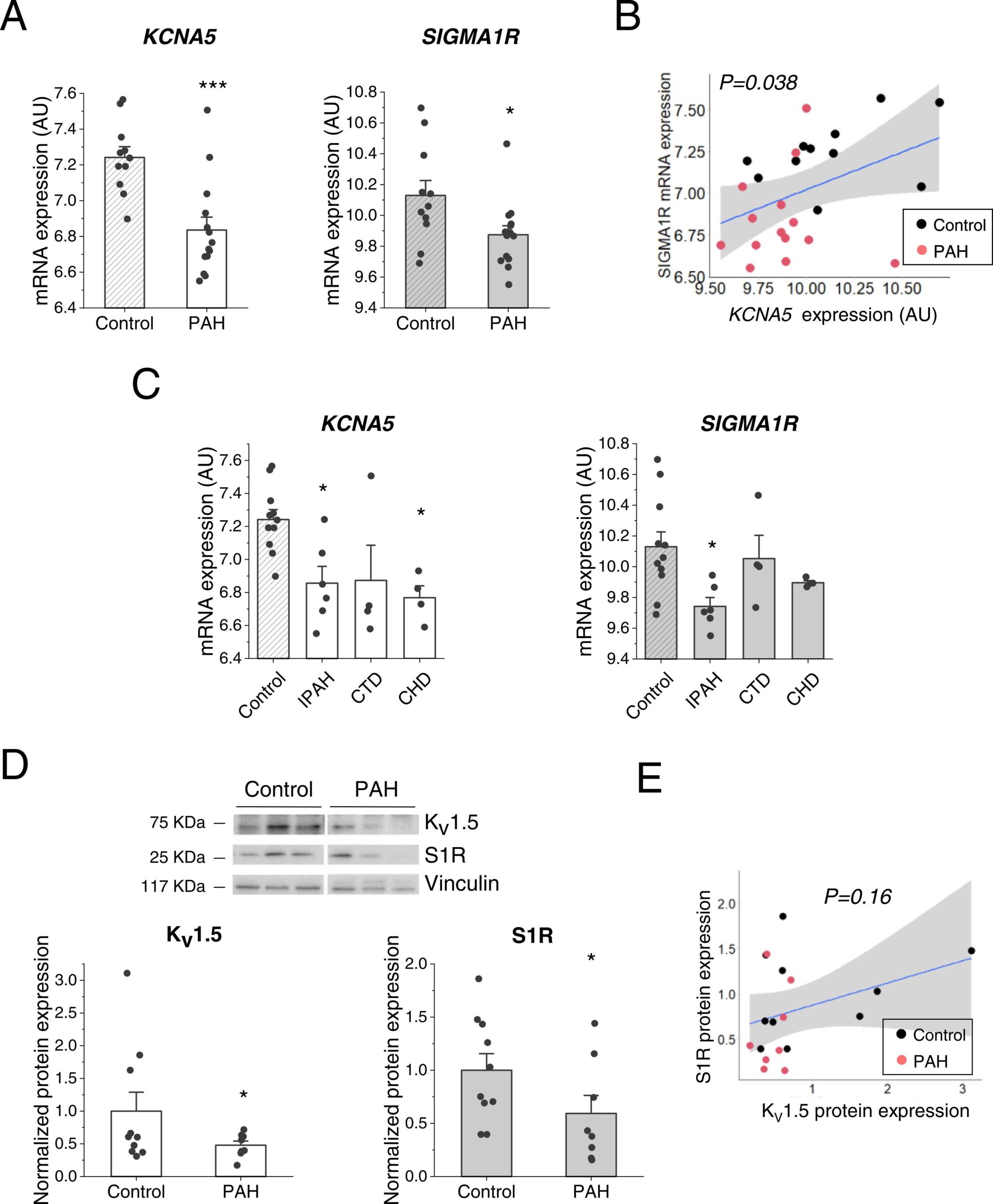
Kv1.5 and S1R mRNA and protein expression in PAH human samples.
(A) mRNA expression levels of KCNA5 (left) and SIGMA1R (right) genes in control (N = 11) and PAH (N = 14) patients after a transcriptomic analysis. (B) Correlation between SIGMA1R and KCNA5 mRNA levels (N = 25). The regression line is in blue, and the 95 % confidence interval is highlighted in grey. P = 0.038. (C) KCNA5 (left) and SIGMA1R (right) mRNA expression dividing the PAH samples between: IPAH (N = 6), CTD (N = 4) and CHD (N = 4). (D) Total KV1.5 (left) and S1R (right) protein expression were quantified in human samples from control (N = 10) or PAH patients (N = 8). Protein expression levels were probed by western blot with an anti-KV1.5 or anti-S1R antibody and normalized by vinculin expression. (E) Correlation between S1R and KV1.5 protein expression (N = 18). The regression line is shown in blue, and the 95 % confidence interval, in grey. P = 0.16. * : P < 0.05, * ** : P < 0.001 vs. control. Data were analysed by a two-tailed (panel A) or one-tailed unpaired student t-test (panel D) or 1-way ANOVA followed by a post-hoc Dunnett test (panel C).
Western-blot analyses also revealed that KV1.5 and S1R were downregulated in AF samples compared with the control ones (Fig. 6A) and both proteins exhibited a positive correlation with a P-value of 0.0008 (Fig. 6B). Additionally, in all types of AF studied: paroxysmal, permanent, and perioperative, S1R was downregulated, but in the case of KV1.5, it was downregulated in permanent and perioperative AF but not in paroxysmal (Fig. 6C).
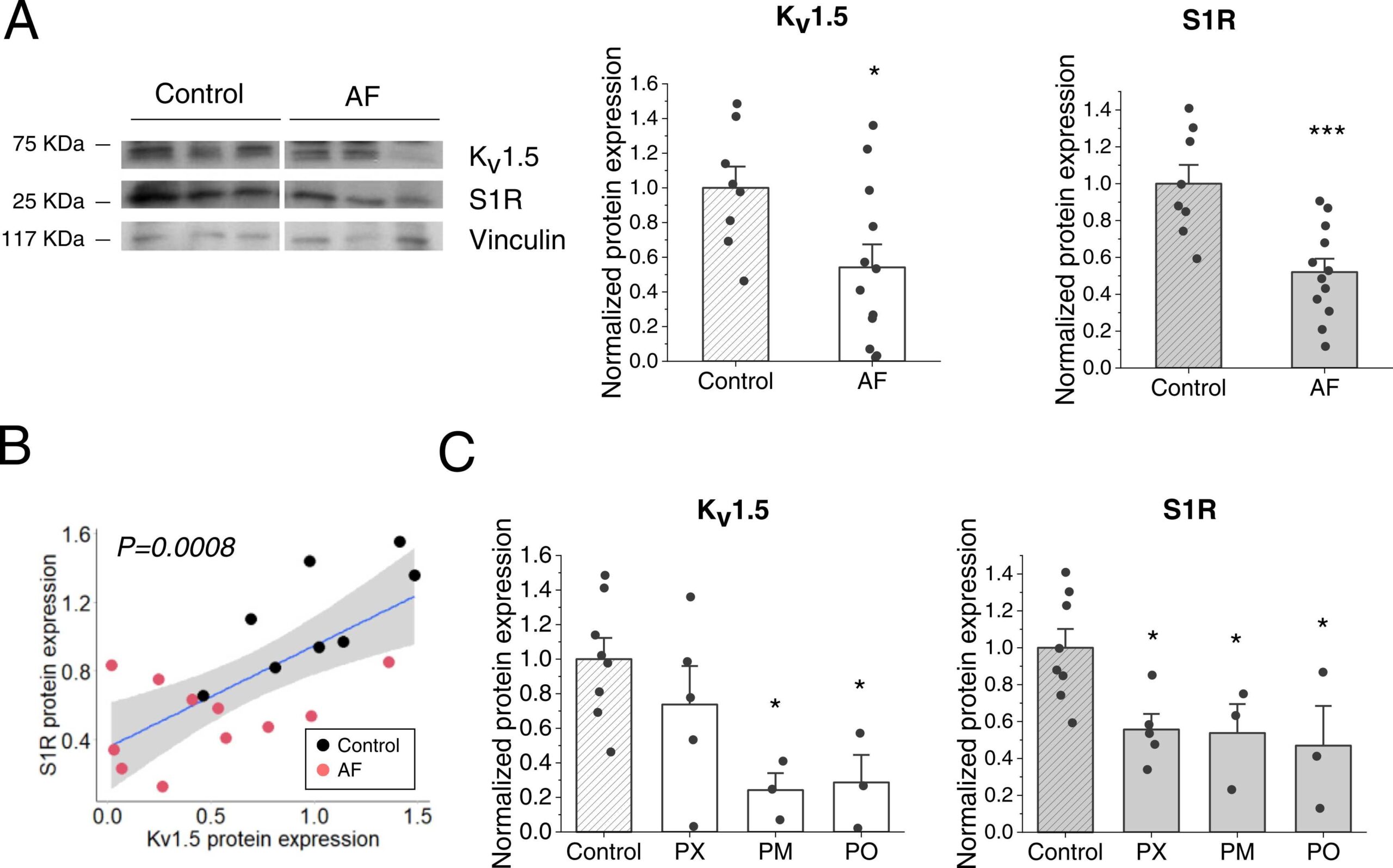
KV1.5 and S1R protein expression in AF human samples.
(A) Total KV1.5 (left) and S1R (right) protein expression was quantified in human samples from control (SR) (N = 8) or AF patients (N = 12). Protein expression levels were probed by western blot with an anti- KV1.5 or anti-S1R antibody and normalized by vinculin expression. (B) Correlation between S1R and KV1.5 protein expression. The regression line is in blue and in grey is indicated the 95 % confidence interval. P = 0.0008 (N = 19). (C) Normalized KV1.5 (left) and S1R (right) protein expression dividing the AF samples between: PX: paroxysmal (N = 5), PM: permanent (N = 3), and PO: perioperative (N = 3). Protein expression levels were normalized to their control. * : P < 0.05, * ** : P < 0.001 vs. control. Data were analysed by a two-tailed unpaired student t-test (panel A) or 1-way ANOVA followed by a post-hoc Dunnett test (panel C).
3.5. The S1R agonist PRE084 preserves KV1.5 channel activity in PASMC under hypoxic conditions
Finally, we studied the potential beneficial effects of PRE084 on KV1.5 channel activity in PA after sustained exposure to hypoxia as an in vitro PAH model. Following 48 h exposure to hypoxia KV currents present in PASMC, which mainly reflect KV1.5 channels as demonstrated by DPO-1 sensitivity, were markedly reduced (Fig. 7A). Notably, treatment with PRE084 was able to markedly increase the attenuated KV currents after hypoxic exposure (Fig. 7A right). The current-voltage relationships of the total KV current and the DPO-1-sensitive (i.e. KV1.5) current are shown in Fig. 7B and C, respectively. Under hypoxic conditions the KV1.5 channel component recorded at membrane potentials more positive than −30 mV were significantly increased by PRE084 treatment (Fig. 7C), reaching values similar to those recorded after incubation under normoxic conditions. Finally, pulmonary arteries were maintained in hypoxia for 48 h with and without PRE084 prior to functional experiments in the myograph. The vasoconstrictor response to KCl was unaffected by incubation with 20 µM PRE084 (2.41 mN/mm2, n = 7 vs. 1.86 mN/mm2, n = 6). On the other hand, the vasoconstriction induced by DPO-1 after hypoxic exposure was greater in PRE084-treated PA than in untreated ones (Fig. 7D). Moreover, PRE084 attenuated the exacerbated contraction induced by serotonin under hypoxic conditions, with no effect on the potency (see Supplementary material online, Table S3). Finally, we determined KV1.5 channel and S1R protein levels to ascertain whether the effects of PRE084 were associated with changes in protein expression. We found that KV1.5 channel expression was not significantly altered by hypoxic exposure, while the addition of PRE084 led to a substantial augmentation of KV1.5 channel expression (seeSupplementary material online, Fig. S7). In contrast, S1R protein levels were comparable in the three groups.
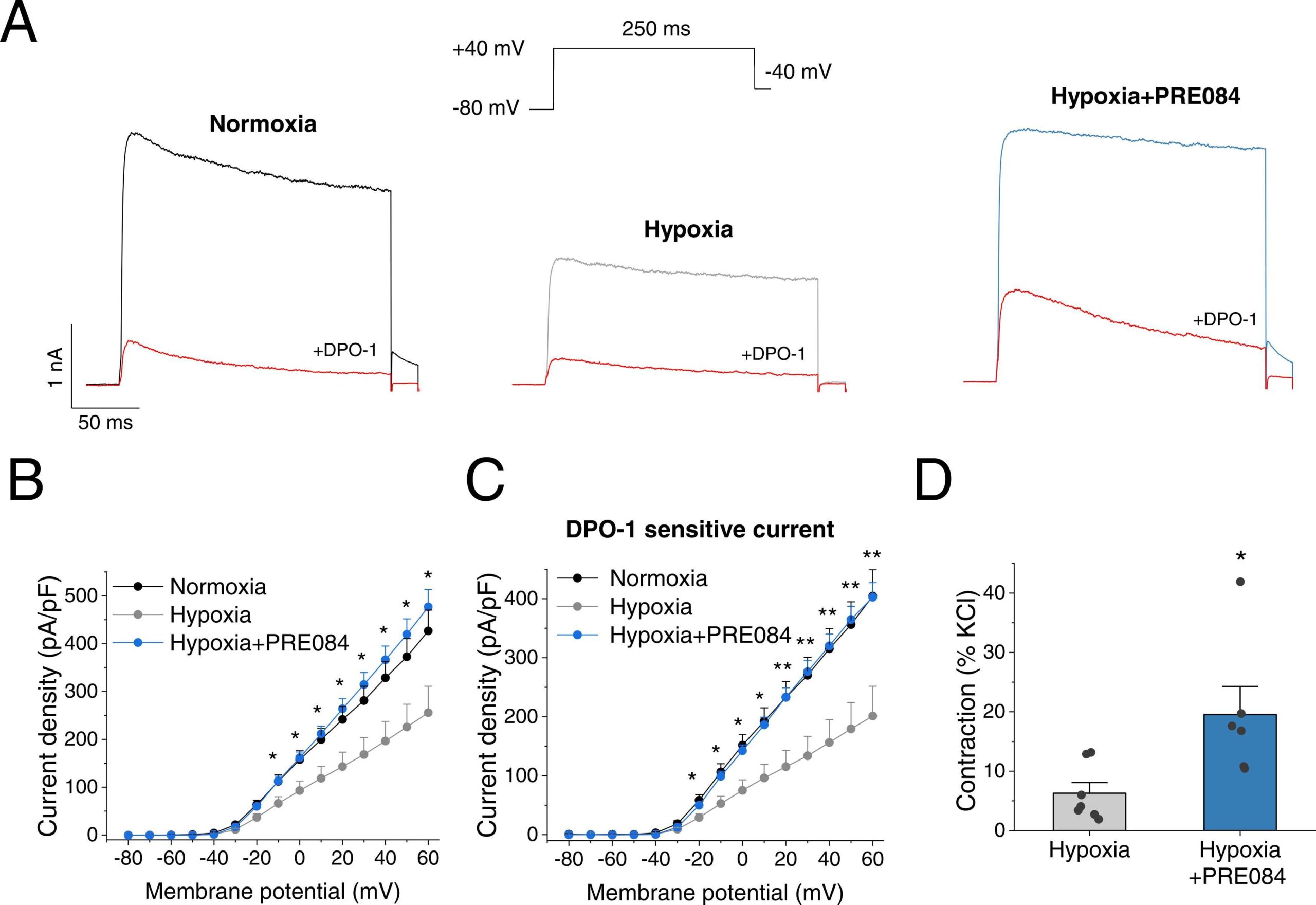
PRE084 increases KV1.5 currents in PASMC following hypoxic exposure.
(A)Representative current traces of K+ currents at + 40 mV in rat PASMC after 48 h incubation under normoxic (left, black), hypoxic (grey) or hypoxic plus PRE084 (20 µM) (right, blue) conditions. The current was recorded in the absence and in the presence of DPO-1 (red). (B) Current-voltage relationships obtained after plotting the maximum current amplitude, normalized by cell capacitance, at each membrane potential (Control n = 9, hypoxia n = 9, hypoxia+PRE0804 n = 6 from N = 6–7 rats per group) and (C) the DPO-1 sensitive current (Control n = 8, hypoxia n = 8, hypoxia+PRE0804 n = 6 from N = 6–7 rats per group). * and * * indicate P < 0.05 and P < 0.01 hypoxia+PRE084 vs hypoxia, respectively. (D) Average data of the vasoconstriction response to DPO-1 in hypoxic-incubated PA untreated or treated with PRE084 (20 µM). (n = 6–7, from N = 6–7 rats per group) * : P < 0.05 hypoxia vs hypoxia+PRE084. Data were analysed by a 1-way ANOVA followed by a post-hoc Dunnett test (panels B and C) or two-tailed unpaired student t-test (panel D). Data are expressed as mean ± SEM.
4. Discussion
In the present study, we have characterized the modulation of KV1.5 channels by S1R. We demonstrate a physical interaction between both proteins. Interestingly, co-transfection with S1R exerts a bimodal regulation of KV1.5 channel expression and activity consisting of up-regulation and down-regulation as S1R levels rise. S1R ligands regulate KV1.5 channel expression and activity in a S1R-dependent manner. Thus, the S1R agonist PRE084 increases KV1.5 current amplitude in PASMC and attenuates pulmonary vasoconstriction and proliferation. Furthermore, KV1.5 and S1R protein expressions are reduced in human heart and lung samples from AF and PAH patients, respectively. Finally, the ability of PRE084 to increase KV1.5 activity is preserved under hypoxic conditions. Our data identifies S1R as a novel regulator of KV1.5 channels and a potential pharmacological target to explore for cardiovascular diseases associated with KV1.5 channel dysregulation.
KV1.5 channels are found throughout the cardiovascular system carrying out essential functions. Thus, KV1.5 channels are key regulators of pulmonary and systemic vascular tone and atrial repolarization [4], [5], [7]. Importantly, KV1.5 expression and activity are altered in different cardiovascular diseases. Thus, downregulation of K+ channels, particularly KV1.5 and TASK-1, is considered an early contributor to the pathophysiology of PAH [4], [6], [30], [31], [32]. Similarly, AF is associated with downregulation of most K+channels, including KV1.5 [33], [34]. Thus, the search for strategies to enhance the activity of KV1.5 channel in these pathological entities has attracted much attention.
S1R regulates the function of a variety of proteins including ion channels, which can occur through direct protein-protein interactions or indirectly through G proteins [9], [35]. Moreover, the physical interaction of S1R with an ion channel may affect its gating, trafficking, maturation or expression [12], [25], [26]. Our data reveal that S1R and KV1.5 channels physically interact and that KV1.5 channel activity and expression are closely linked to S1R levels. Thus, S1R causes a bimodal modulation of KV1.5 channels in both cell types, characterised by an increase and a decrease in activity and expression as S1R levels increase. Aydar E et al. [10] found a similar depression of KV1.4 currents in Xenopus oocytes coexpressed with a 1:1 S1R:Kv1.4 ratio or higher but these authors did not performed experiments overexpressing lower amounts of S1R. Apart from current amplitude, S1R did not significantly influenced voltage-dependent activation and inactivation parameters of KV1.5 channels hence strongly suggesting that S1R mainly affects their expression rather than their gating. Unlike KV1.5, KV2.1 or Kir2.1 currents were not affected by S1R overexpression. Our data on KV2.1 are consistent with the negligible effect of S1R knockdown previously reported [36] while the lack of effect on Kir2.1 is not in agreement with the proposed S1R-Kir2.1 functional coupling [37]. All these data demonstrate that S1R could represent a new member of the KV1.5 channelosome [38].
We next explored the effects of S1R ligands on KV1.5 channels. Two previous studies have evaluated the effect of S1R agonists on KV1.5 currents in heterologous systems [10], [26]. Aydar et al. reported that in Xenopus oocytes transfected with KV1.5 and S1R, the S1R agonist SKF10047 (100 µM) acutely perfused (2 min) inhibited KV1.5 current in a S1R-dependent manner [10]. A more recent study found a similar inhibitory effect on KV1.5 by PRE084 and SKF10047 in HEK293 cells [26]. Herein we also observed that acute administration of these S1R ligands led to a substantial inhibition of the KV1.5 current. In line with previous studies on KV1.2 [26], KV1.3 [13], and KV2.1 [36] we found that the inhibitory effect of S1R ligands was unaffected when S1R was co-expressed. Moreover, we observed a similar inhibitory effect when testing the S1R antagonist BD1047. These data challenges whether the inhibitory effects observed after acute exposure to S1R ligands are indeed mediated via S1R.
Since our data suggested that KV1.5 regulation by S1R involves changes in channel expression we examined the chronic effects of S1R ligands on KV1.5 current and expression. We found that 24 h exposure to SKF10047 and PRE084 led to a substantial increase in KV1.5 current which was associated with an increased expression of the channel. Importantly, these effects were observed when HEK293 cells were co-expressed with KV1.5 and S1R but not when cells were only transfected with the channel, indicating that KV1.5 upregulation by the ligands is S1R dependent. Although studies analysing the chronic effect of S1R ligands on KV1.5 channels are lacking, there is evidence that other KV channels can be upregulated in an S1R-dependent manner [12]. Conversely, KV1.5 channel expression was decreased by the S1R antagonist BD1047. In addition, S1R protein levels in HEK293 cells and isolated PA were not altered by S1R ligands, which suggest that these ligands affect the oligomerization state of S1R rather than its expression. Further research is needed to elucidate the precise functional consequences of S1R oligomerization, but it seems to be key for the interaction with target proteins [14].
Overall, our data showed that KV1.5 channel activity and expression may be fine-tuned by S1R ligands and encouraged us to move to a more physiological system. Thus, we evaluated the effects of PRE084 in pulmonary arterial myocytes characterised by high KV1.5 channel activity [30]. We found that 24 h incubation with PRE084 produced a strong increase in the KV1.5 current present in PASMC. In the vascular system, an increase in the KV1.5 current limits membrane depolarization and thus reduces the open probability of the voltage-gated calcium channels, hence decreasing Ca2+ entry and attenuating vasoconstriction [4], [5], [30]. In this sense, our vascular reactivity experiments showed that PRE084-exposed PA had an attenuated contraction to Phe in a concentration-dependent manner. Importantly, this effect was prevented in the presence of the KV1.5 channel inhibitor DPO-1, which strongly suggests the increase in KV1.5 channel activity as the underlying mechanism for the attenuated vasoconstriction.
In addition to vascular tone regulation, KV1.5 channel activity controls cellular processes such as apoptosis and proliferation, being an increase in the KV1.5 current associated with a decrease in early and late apoptosis in PASMC [2]. In this regard, K+ is the dominant cation in the cytoplasm and thus plays an important role in the maintaining of the cell volume. In early apoptosis, decreased KV activity inhibits apoptotic cell shrinkage, and in later stages of apoptosis, decreases caspase activation and DNA fragmentation [2], [39]. Furthermore, S1R plays an important role in counteracting ER stress and decreasing apoptosis [8], [20]. In fact, PRE084 has also been shown to reduce cell apoptosis [40]. Here, we demonstrate that PRE084 decreases PASMC proliferation in a concentration-dependent manner. Nevertheless, our findings do not support an effect of PRE084 on apoptosis in PASMC.
Considering the ability of S1R agonists to modulate KV1.5 channels and affect pulmonary vascular function and given the role of KV1.5 channel dysfunction in PAH, we also examined the S1R expression in human PAH samples. A transcriptomic analysis revealed decreased KV1.5 mRNA in lung samples from PAH samples which was concurrent with diminished S1R. In line with these results, protein levels of both KV1.5 and S1R were also decreased in PAH samples. Likewise, a similar protein downregulation of KV1.5 and S1R was observed in cardiac human samples from AF, in which KV1.5 channel dysfunction is also considered a key contributor. Consistent with these data, several experimental studies have also found a reduced S1R expression in the atrium of a rat model of AF [22]and in the right ventricle of PAH rats [41]. In our study mRNA and protein expression of KV1.5 and S1R showed a positive correlation in both pathologies. Nevertheless, further research is needed to elucidate whether the S1R downregulation has any causal relationship with lower KV1.5 expression in AF and/or PAH and could contribute to the pathophysiology of these diseases. Moreover, studies in animal models of these diseases would be necessary to determine whether the reduced expression of S1R may limit the therapeutic potential of S1R agonists in these pathological conditions.
We finally evaluated the potential beneficial effects of PRE084 on KV1.5 channel dysfunction following sustained hypoxic exposure, which represents a standard in vitro model of PH due to its role as an underlying trigger for different forms of pulmonary hypertension including PAH (group I PH) and especially that due to respiratory diseases (group III PH). Interestingly, we found that PRE084 was able to rescue hypoxic downregulation of KV1.5 channel activity and to attenuate the exacerbated 5-HT contraction observed after hypoxic exposure, pointing to a potential benefit effect in PH associated with hypoxia. Of note, the reduced KV1.5 channel activity in PA after 48 h hypoxic exposure was not associated with changes in KV1.5 channel or S1R protein expression. Previous studies have shown that in PASMC, mRNA levels of KV1.5 are reduced after 48 h incubation with hypoxia [42], while the reduction in the amount of KV1.5 channel protein seems to require longer incubations (≥60 h) [42], [43], [44]. This suggests that the attenuated KV1.5 activity observed after ≤ 48 h of hypoxia observed herein and previously reported [31], [45], [46] does not appear to be strictly related to a decrease in the total amount of KV1.5 channel protein but may involve, among others, alterations in gating, trafficking, or surface expression of the channel [4], [5].
The ability of PRE084 to increase KV1.5 current under both normoxic and hypoxic conditions may be of great relevance. Since KV1.5 channels are considered a key mechanism for hypoxic pulmonary vasoconstriction [28], [43], [47], [48], [49], their preservation by PRE084 under hypoxic conditions is expected to maintain this crucial protective mechanism that divert blood flow away from hypoxic lung areas to the best oxygenated alveoli [49]. Although more research is needed to fully address these questions, there are several examples of protective effects of S1R agonists such as PRE084 in the cardiovascular system, while the antagonists produce the opposite effects [40], [50], [51]. Moreover, Sun et al. has recently shown that activation of S1R improves right ventricular dysfunction secondary to pulmonary hypertension [41]. Intriguingly, several therapeutic drugs with S1R agonistic properties such as donepezil, the serotonin reuptake inhibitors fluvoxamine and fluoxetine, or dehydroepiandrosterone have shown beneficial effects in experimental PH [41], [52], [53], [54]. Moreover, fluoxetine-induced protection was linked to KV1.5 channels upregulation [53], [55]. Whether S1R activation contributes to these effects requires further studies.
In conclusion, our data reveal that S1R functionally interacts with the KV1.5 channel and acts as a critical regulator of its expression and activity. In the arterial pulmonary vessels, PRE084 is able to increase KV1.5 currents and reduce contraction and proliferation of PASMC. Of note S1R expression is downregulated in human AF and PAH. Altogether, these data identify S1R as a potential novel pharmacological target in diseases associated with an impaired KV1.5 channel function.
5. Conclusions
Pulmonary arterial hypertension and atrial fibrillation are severe cardiovascular diseases associated with electrical remodelling including KV1.5 channel dysfunction. Thus, novel pharmacological strategies to improve the attenuated KV1.5 channel function are highly demanded. Here, we show that the chaperone S1R finetunes KV1.5 channel expression and function and S1R modulators may effectively enhance KV1.5 channel function. Our study identifies S1R as a novel pharmacological target to rescue the attenuated KV1.5 channel function in cardiovascular diseases associated with channel malfunction.