
Gut microbiota from sigma-1 receptor knockout mice induces depression-like behaviors and modulates the cAMP/CREB/BDNF signaling pathway
By Jia-Hao Li, Jia-Li Liu, Xiu-Wen Li, Yi Liu, Jian-Zheng Yang, Li-Jian Chen, Kai-Kai Zhang, Xiao-Li Xie, and Qi Wang
Excerpt from the article published in Frontiers in Microbiology, 06 April 2023, Sec. Evolutionary and Genomic Microbiology, Volume 14 – 2023, DOI: https://doi.org/10.3389/fmicb.2023.1143648
Editor’s Highlights
- In the past few decades, various theories about the development of depression Depression, also known as major depressive disorder (MDD), have been proposed, including the neurotrophic hypothesis.
- The neurotrophic hypothesis suggests that brain-derived neurotrophic factor (BDNF) and other neurotrophic factors may be involved in the development of depression, and reductions in BDNF have been linked to atrophy of brain regions involved in emotion, such as the hippocampus.
- Activation of Sig-1R enhanced the conversion of pro-BDNF to mature BDNF, as well as the release of mature BDNF into the extracellular space. These pieces of evidence suggest that activation of the Sig-1R promotes chaperone activity, which influences BDNF secretion, exerting antidepressant-like effects.
- It is well known that the expression of BDNF is regulated by cAMP (Cyclic adenosine 3′,5′-monophosphate) response element binding protein (CREB).
- The gut microbiota may regulate the expression of neurotrophic factors through the cAMP/CREB/BDNF signaling pathway.
- The microbiota from Sig-1R knockout mice was sufficient to cause a depression-like phenotype and a reduction in neurotrophic factors.
- This study offers a new perspective on the mechanism of action of Sig-1R in the regulation of depression and provides further evidence for the study of the gut-brain axis.
Abstract
Introduction: Depression is a common mental disorder that affects approximately 350 million people worldwide. Much remains unknown about the molecular mechanisms underlying this complex disorder. Sigma-1 receptor (Sig-1R) is expressed at high levels in the central nervous system. Increasing evidence has demonstrated a close association between the Sig-1R and depression. Recently, research has suggested that the gut microbiota may play a crucial role in the development of depression.
Methods: Male Sig-1R knockout (Sig-1R KO) and wild-type (WT) mice were used for this study. All transgenic mice were of a pure C57BL/6J background. Mice received a daily gavage of vancomycin (100 mg/kg), neomycin sulfate (200 mg/kg), metronidazole (200 mg/kg), and ampicillin (200 mg/kg) for one week to deplete gut microbiota. Fecal microbiota transplantation (FMT) was conducted to assess the effects of gut microbiota. Depression-like behaviors was evaluated by tail suspension test (TST), forced swimming test (FST) and sucrose preference test (SPT). Gut microbiota was analyzed by 16s rRNA and hippocampal transcriptome changes were assessed by RNA-seq.
Results: We found that Sig-1R knockout induced depression-like behaviors in mice, including a significant reduction in immobility time and an increase in latency to immobility in the FST and TST, which was reversed upon clearance of gut microbiota with antibiotic treatment. Sig-1R knockout significantly altered the composition of the gut microbiota. At the genus level, the abundance of Alistipes, Alloprevotella, and Lleibacterium decreased significantly. Gut microbiota dysfunction and depression-like phenotypes in Sig-1R knockout mice could be reproduced through FMT experiments. Additionally, hippocampal RNA sequencing identified multiple KEGG pathways that are associated with depression. We also discovered that the cAMP/CREB/BDNF signaling pathway is inhibited in the Sig-1R KO group along with lower expression of neurotrophic factors including CTNF, TGF-α and NGF. Fecal bacteria transplantation from Sig-1R KO mice also inhibited cAMP/CREB/BDNF signaling pathway.
Discussion: In our study, we found that the gut-brain axis may be a potential mechanism through which Sig-1R regulates depression-like behaviors. Our study provides new insights into the mechanisms by which Sig-1R regulates depression and further supports the concept of the gut-brain axis.
1. Introduction
Depression, also known as major depressive disorder (MDD), is a common mental disorder that affects approximately 350 million people worldwide. The causes of depression are thought to be multifaceted and may involve both genetic and environmental factors (Malhi and Mann, 2018). In the past few decades, various theories about the development of depression have been proposed, including the inflammation hypothesis, the neural circuits hypothesis, the neurotransmitter hypothesis, and the neurotrophic hypothesis (López-Figueroa et al., 2004; Duman and Monteggia, 2006; Fujimoto et al., 2008; Maes, 2008). The neurotrophic hypothesis suggests that brain-derived neurotrophic factor (BDNF) and other neurotrophic factors may be involved in the development of depression, and reductions in BDNF have been linked to atrophy of brain regions involved in emotion, such as the hippocampus (Castren and Rantamaki, 2010). Neurotrophic factor BDNF has also turned out to be significantly associated with depression in clinical patients and in extensive studies. In recent years, the neurotrophic hypothesis has become an important target of novel antidepressant drugs (Wang A. et al., 2021). Despite significant progress in our understanding of depression, much remains unknown about the molecular mechanisms underlying this complex disorder.
Sigma-1 receptor is a protein encoded by the SIGMAR1 gene. Sig-1R is expressed at high levels in the central nervous system and has been shown to have neuroprotective effects (Wu et al., 2021). Mutations in the SIGMAR1 gene have been linked to an increased risk of cognitive dysfunction and are strongly associated with Alzheimer’s disease (AD; Fehér et al., 2012). In addition, two studies have found that Sig-1R may be involved in the development of depression through its effects on neurotrophic and growth factor signaling pathways (Kishi et al., 2010; Mandelli et al., 2017; Stiernstromer et al., 2022). Selective serotonin reuptake inhibitors (SSRIs) have a moderate affinity for Sig-1Rs and the antidepressant effect of SSRIs may be mediated by Sig-1R (Smith and Su, 2017; Izumi et al., 2023). Efficacy of fluvoxamine treatment in improving psychotic symptoms and depressive symptoms is dose-dependently related to its binding to Sig-1Rs in the healthy human brain (Hashimoto, 2009). One potential mechanism through which Sig-1R may exert its neuroprotective effects is by promoting brain-derived neurotrophic factor (BDNF) expression. For example, Sig-1R agonists have been shown to reverse the downregulation of BDNF and improve symptoms of post-traumatic stress disorder (PTSD; Ji et al., 2017). Another Sig-1R agonist has been demonstrated to have neuroprotective effects in a mouse model of alpha-thalassemia X-linked intellectual disability (Yamaguchi et al., 2018). The brain’s BDNF protein was elevated by chronic antidepressant treatment, including SSRIs (Hashimoto, 2010). Activation of Sig-1R enhanced the conversion of pro-BDNF to mature BDNF, as well as the release of mature BDNF into the extracellular space (Fujimoto et al., 2012; Hashimoto, 2013). These pieces of evidence suggest that activation of the Sig-1R promotes chaperone activity, which influences BDNF secretion, exerting antidepressant-like effects. Despite these advances, the exact role of Sig-1R in the development of depression is not yet fully understood.
The relationship between gut microbiota and depression has garnered increasing attention in recent years, with a growing body of evidence demonstrating a significant association between the two. Studies have found that the gut microbiota of individuals with depression differs significantly from that of healthy individuals (Jiang et al., 2015; Aizawa et al., 2016). Moreover, transplanting the microbiota from depressed individuals into germ-free or antibiotic-treated mice has resulted in the induction of symptoms similar to depression (Kelly et al., 2016; Zheng et al., 2016; Li et al., 2019). These findings suggest that the gut microbiota may play a role in the pathogenesis of depression.
Despite the growing evidence linking Sig-1R to depression, the precise role of Sig-1R in this disorder remains unknown. To further understand the involvement of Sig-1R in depression, we conducted a study in which we compared depression-like behaviors in the wild-type (WT) and Sig-1R knockout (KO) groups. We also explored the role of gut microbiota in these behaviors through antibiotic treatment and fecal microbiota transplantation (FMT). Our results showed that Sig-1R knockout mice exhibited increased depression-like behaviors and significant changes in gut microbiota. The gut microbiota of Sig-1R knockout mice was sufficient to induce an increase in depression-like behaviors. These effects may be mediated by a reduction in neurotrophic factors through the cAMP/CREB/BDNF signaling pathway. Our study provides new insights into the mechanisms by which Sig-1R may contribute to the development of depression.
…
3. Results
3.1. Antibiotic treatment reverses sig-1R knockout-induced depression-like behaviors
The ABX treatment flowchart is shown in Figure 1A. In the TST (Figure 1B), Sig-1R knockout mice exhibited a significantly shortened latency to immobility compared with WT mice, and displayed significantly increased immobility time (Figure 1C). This trend was consistent in the FST, where Sig-1R knockout mice displayed a shortened latency to immobility and prolonged immobility time (Figures 1D,E). Additionally, in the SPT, we observed a significant reduction in the sucrose preference index of the KO group compared to that of WT group (Figure 1F). Taken together, these results suggest that Sig-1R knockout elicits depression-like behaviors in mice.
To investigate the potential role of gut microbiota in Sig-1R knockout-induced depression-like behaviors, we administered an antibiotic treatment to clear the gut microbiota in mice. The results of behavioral tests indicated that this treatment eliminated the depression-like behaviors in Sig-1R knockout mice, as evidenced by the lack of significant differences in latency to immobility, immobility time, and sucrose preference index between the WT + ABX and KO + ABX groups in the TST, FST, and SPT experiments (Figures 1B–F). These findings suggest that gut microbiota may contribute to the development of Sig-1R knockout-mediated depression-like behaviors, and that antibiotic treatment can effectively eliminate these behaviors.
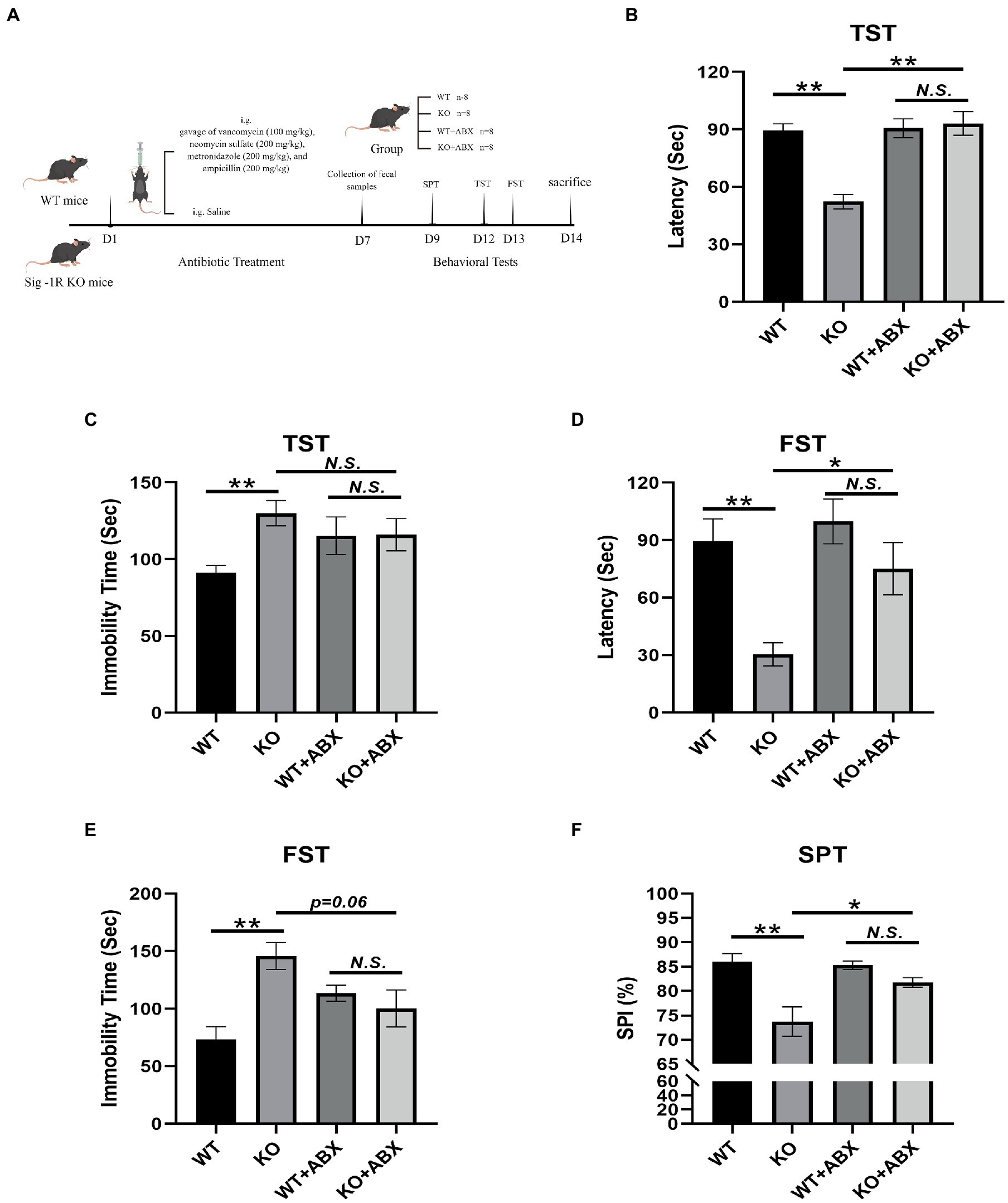
Antibiotic treatment reverses Sig-1R knockout-induced depression-like behaviors.
(A)The ABX treatment flowchart by Figdraw. (B)Latency (s) to immobility in the TST [For ABX treatment, F(1, 13) = 11.50, p = 0.0048, two-way ANOVA]. (C) Immobility time (s) in the TST [For ABX treatment, F(1, 13) = 6.068, p = 0.0285, two-way ANOVA]. (D) Latency (Sec) to immobility in the FST [For ABX treatment, F(1, 13) = 5.950, p = 0.0298, two-way ANOVA]. (E) Immobility time (s) in the FST [For ABX treatment, F(1, 13) = 13.37, p = 0.0029, two-way ANOVA]. (F) The sucrose preference index (SPI) in the SPT [For ABX treatment, F(1, 10) = 37.09, p = 0.0001, two-way ANOVA]. **p < 0.01. *p < 0.05. N.S.: not significant. WT, n = 8; KO, n = 8; WT + ABX, n = 6; and KO + ABX, n = 6.
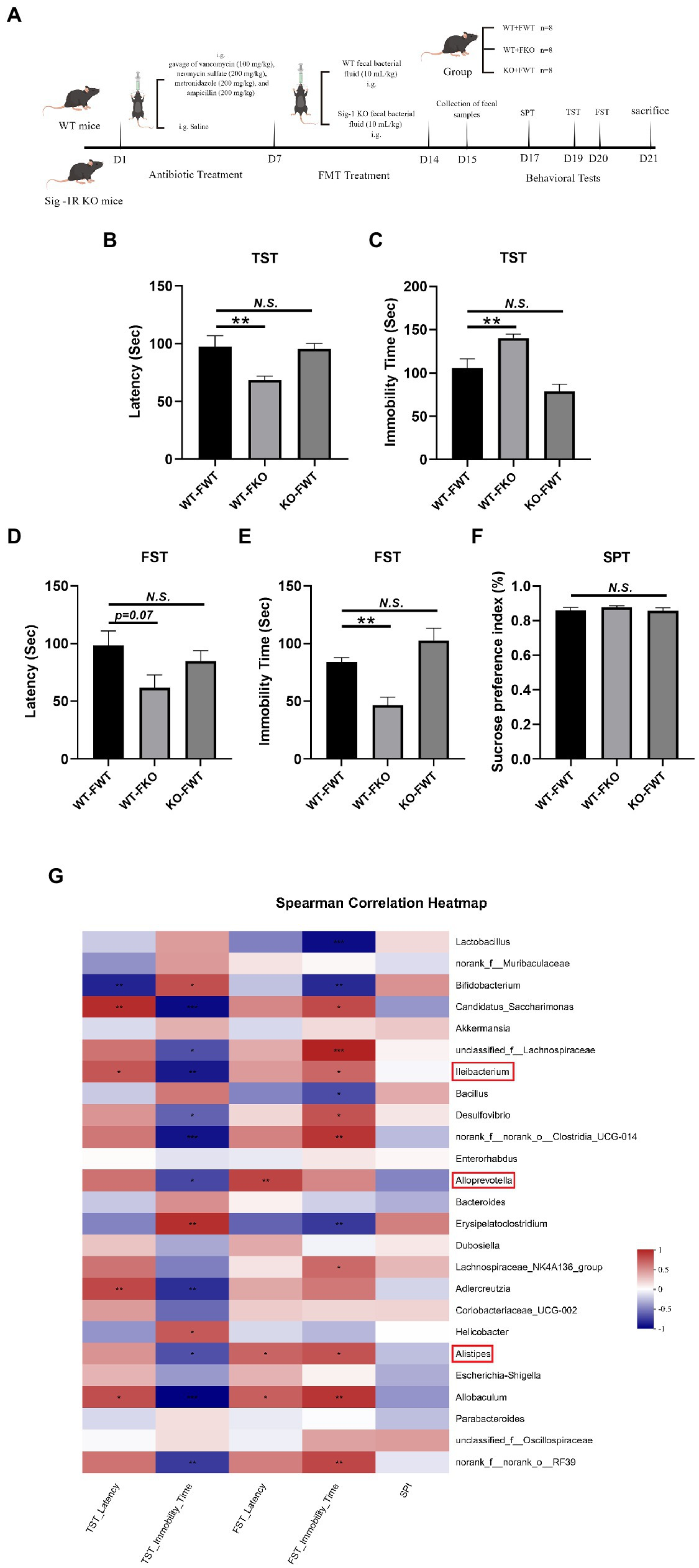
Sig-1R KO gut microbiota induced depression-like behaviors.
(A) The FMT treatment flowchart. By Figdraw. (B) Latency (s) to immobility in the TST. F(2, 18) = 6.048, p = 0.0098. (C) Immobility time (s) in the TST. F(2, 18) = 14.20, p = 0.0002. (D) Latency (s) to immobility in the FST. F(2, 15) = 2.884, p = 0.0871. (E) Immobility time (s) in the FST. F(2, 15) = 13.60, p = 0.0004. (F)The sucrose preference index (SPI) in the SPT. F(2, 15) = 0.6173, p = 0.5523. **p < 0.05. N.S.: not significant. WT-FWT, n = 7; WT-FKO, n = 7; and KO-FWT, n = 7. (G) The correlation between the gut microbiota and depressive-like behaviors in recipient mice was analyzed using Spearman’s correlation analysis. The red color represents a positive correlation; the blue color represents a negative correlation. Asterisk-marked columns indicated significant correlation. *0.01 < p ≤ 0.05, **0.001 < p ≤ 0.01, and ***p ≤ 0.001.
3.2. Sig-1R knockout induced dysbiosis of the gut microbiota
To examine the impact of Sig-1R knockout on gut microbiota composition, we conducted 16S rRNA sequencing and analyzed alpha diversity using the Ace index (Figure 3A) and Simpson index (Figure 3B). While we did not observe significant differences in alpha diversity between the KO and WT groups, principal coordinate analysis (PCoA) of beta diversity revealed clear differences in microbiota profiles between the two groups, with samples from each group clustering together (Figure 3C). Hierarchical clustering analysis also indicated significant differences in gut microbiota between the WT and KO groups (Figure 3D). Linear discriminant analysis effect size (LEfSe) analysis revealed significant differences in bacterial abundance at various levels of taxonomic classification, from the phylum to genus level (Figure 3E). Using the Wilcoxon rank-sum test, we further analyzed differences in gut microbiota at the genus level (Figure 3F) and found that the relative abundance of Alistipes, Alloprevotella, and Lleibacterium was significantly decreased in the KO group compared to the WT group (Figures 3G–I). We conducted correlation analysis to investigate the relationship between depression-like behaviors and changes in the gut microbiota, and found that the gut microbiota was closely related to depression-like behaviors (Figure 3J). These results suggest that Sig-1R knockout leads to dysbiosis of the gut microbiota.
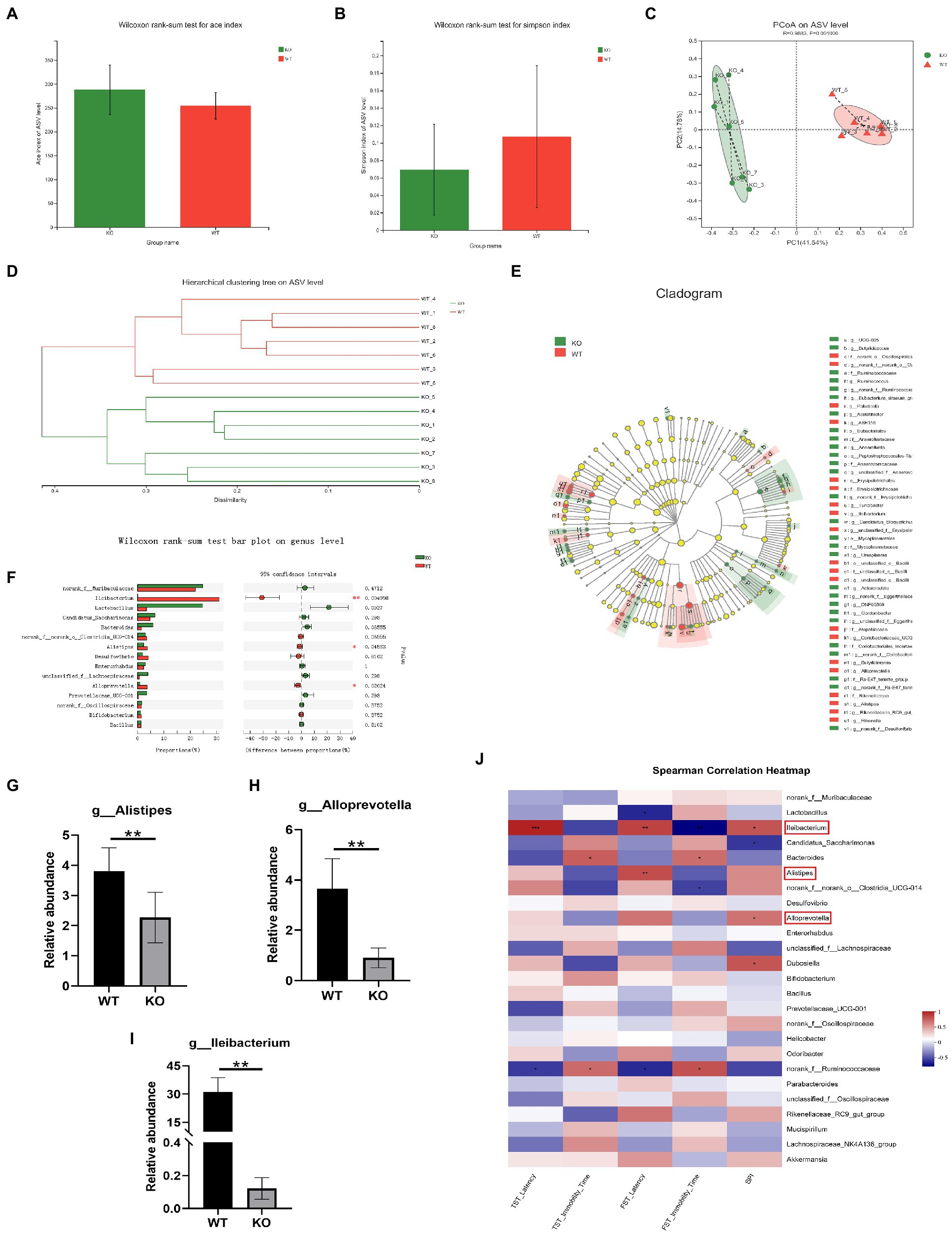
Sig-1R knockout induced dysbiosis of the gut microbiota.
(A) ACE index and (B) Simpson index of the gut microbiota. (C) PCoA and (D)hierarchical clustering of 16S rRNA gene sequences. (E) The LEfSe analysis of the gut microbiota from the phylum level down to the genus level. (F) Composition of the gut microbiota at the genus level. (G–I) The relative abundance of Alistipes, Alloprevotella, and Lleibacterium. **p<0.01. WT, n = 6; KO, n = 6. (J)The correlation between the gut microbiota and depressive-like behaviors in WT and KO mice was analyzed using Spearman’s correlation analysis. The red color represents a positive correlation; the blue color represents a negative correlation. Asterisk-marked columns indicated significant correlation. *0.01 < p ≤ 0.05, **0.001 < p ≤ 0.01, and ***p ≤ 0.001.
3.3. FMT alters the gut microbiota in recipient mice
To examine the relationship between gut microbiota and depression-like behaviors, we performed an FMT experiment in which the gut microbiota of KO or WT groups were transplanted into recipient mice. The flowchart of the modeling process is shown in Figure 2A. Using 16S rRNA sequencing, we found that the microbiota of the recipient mice resembled that of their respective donor mice (Figure 4A). We then compared the alpha diversity of the WT-FWT and WT-FKO groups. The Ace index (Figure 4B) and Simpson index (Figure 4C) showed that the WT-FKO group had a significantly lower diversity and richness of bacterial species compared to the WT-FWT group. Beta diversity analysis using PCoA (Figure 4D) and hierarchical clustering (Figure 4E) revealed significant differences in the composition of the two groups, with samples from each group clustering together. LEfSe analysis showed differential gut microbiota at various levels of taxonomic classification, from the phylum to genus level (Figure 4F). Further analysis at the genus level using the Wilcoxon rank-sum test revealed significant decreases in the relative abundance of Alistipes, Alloprevotella, and Lleibacterium in the WT-FKO group compared to the WT-FWT group (Figures 4H–J). These findings suggest that FMT reproduces the dysbiosis of the gut microbiota observed in Sig-1R knockout mice.
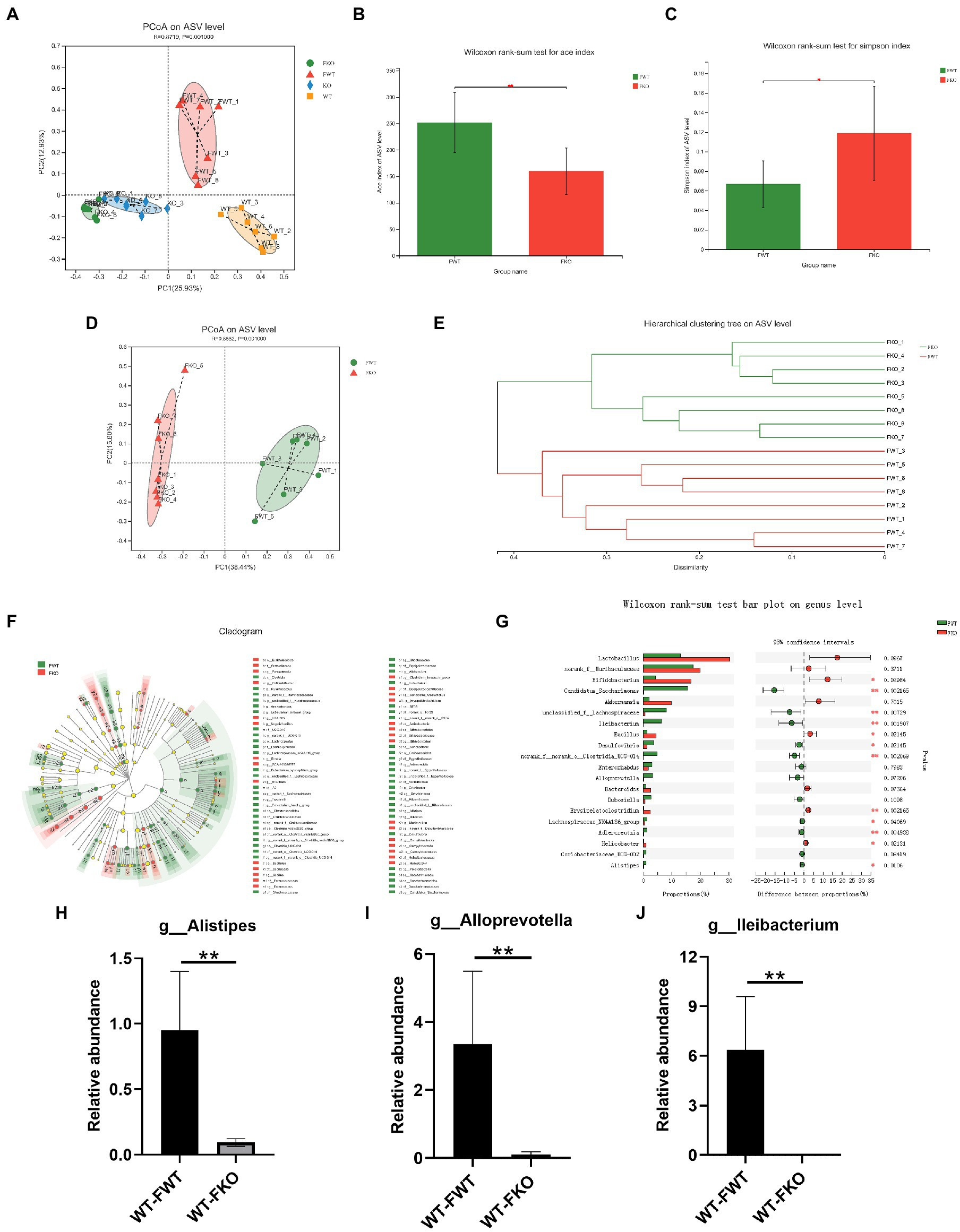
FMT alters the gut microbiota in recipient mice.
(A) The gut microbiota PCoA analysis of four groups of mice. (B) ACE index and (C) Simpson index of the gut microbiota. (D)PCoA and (E) hierarchical clustering of 16S rRNA gene sequences. (F) The LEfSe analysis of the gut microbiota from the phylum level down to the genus level. (G) Composition of the gut microbiota at the genus level. (H–J) The relative abundance of Alistipes, Alloprevotella, and Lleibacterium. **p < 0.01. WT-FWT, n = 6; WT-FKO, n = 6.
3.4. Sig-1R KO gut microbiota induced depression-like behaviors
We performed behavioral tests on FMT recipient mice. The results of the TST and FST experiments showed that the WT-FKO group exhibited a shorter latency to immobility (Figures 2B,D) and increased immobility time (Figures 2C,E) compared to the WT-FWT group, suggesting that feces from Sig-1R knockout mice induced depression-like behaviors in the recipient mice. However, there was no significant difference in SPI between the two groups in the SPT experiment (Figure 2F). In contrast, there were no significant differences in behavioral results between the KO-FWT group and the WT-FWT group, suggesting that feces from WT mice reversed Sig-1R knockout-induced depression-like behaviors. Again, we conducted correlation analysis to investigate the relationship between depression-like behaviors and changes in the gut microbiota, and found that the gut microbiota such as Alistipes, Alloprevotella, and Lleibacterium still showed close correlation with depression-like behavior (Figure 2G).
3.5. cAMP/CREB/BDNF signaling mediated depression-like behaviors in sig-1R knockout mice
To further investigate the mechanism underlying depression-like behaviors in Sig-1R knockout mice, we performed RNA sequencing on the hippocampus of WT and Sig-1R knockout mice. PCA revealed significant differences between the two groups (Figure 5A). Using a fold change threshold of 2 and a value of p of <0.05, a Volcano plot analysis showed that 241 mRNAs were upregulated and 204 were downregulated (Figure 5B). We conducted KEGG enrichment analysis and protein–protein interaction (PPI) analysis on the differentially expressed genes. KEGG enrichment analysis revealed that several pathways related to depression, including the retrograde endocannabinoid signaling pathway, dopaminergic synapse, GABAergic synapse, and cAMP signaling pathway, were enriched (Figure 5C). PPI analysis identified Grin1 and Gria1 as the most central genes in the PPI network; these genes have been previously linked to depression (Figure 5D).
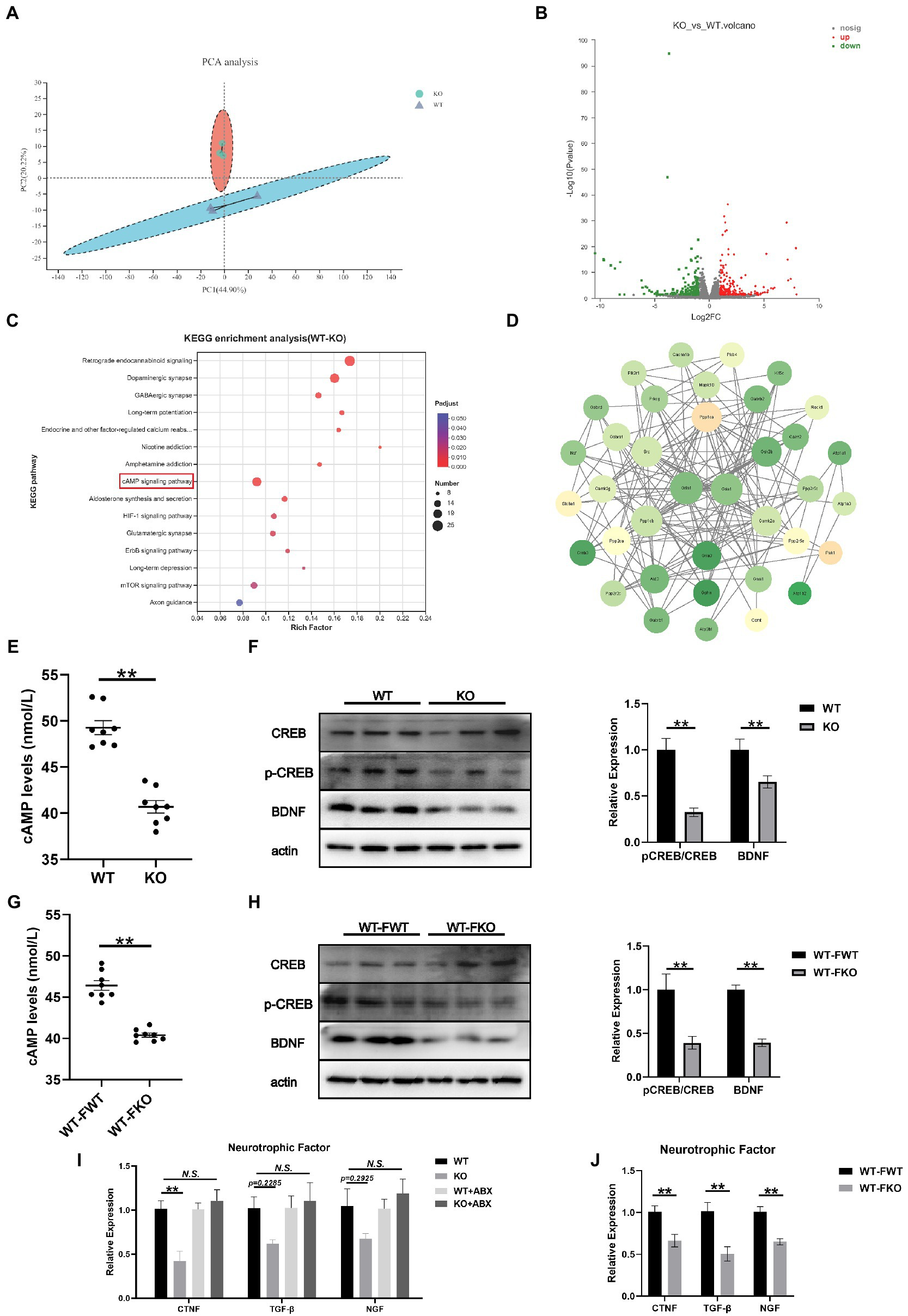
cAMP/CREB/BDNF signaling mediated depression-like behaviors in Sig-1R knockout mice.
(A) PCA of the RNA-seq data. (B) Volcano plots of the DEGs from RNA-seq, with 241 upregulated and 204 downregulated. Red color intensity signifies upregulation of gene expression, and green signifies downregulation. (C) Mapping of the RNA-seq data to the KEGG pathway (KEGG). The vertical axis represents the pathway names, while the horizontal axis represents the enrichment factor. The size of the point indicates the number of DEGs in the pathway, and the color of the point corresponds to a different Q-value range. (D) PPI network analysis. The darker color indicates a higher core degree. (E) cAMP levels in WT and KO group. (F)Western blotting and statistical analysis of cAMP/CREB/BDNF signaling pathway in WT and KO group. (G) cAMP levels in FMT recipient mice. (H) Western blotting and statistical analysis of cAMP/CREB/BDNF signaling pathway in FMT recipient mice. (I,J) RT-qPCR for the expression of neurotrophic factors CTNF, TGF-β, and NGF [CTNF F(3, 12) = 9.073, p = 0.0021; TGF-β F(2, 15) = 2.413, p = 0.1174. F(2, 15) = 2.395, p = 0.1192. One-way ANOVA]. **p < 0.05. N.S.: not significant.
Hippocampus-derived brain-derived neurotrophic factor (BDNF) has been identified as a significant factor in the development of depression. It is well known that the expression of BDNF is regulated by cAMP response element binding protein (CREB). In this study, we aimed to investigate the role of the cAMP/CREB/BDNF signaling pathway in Sig-1R knockout-induced depression-like behaviors. To do this, we compared the levels of cAMP and the expressions of pCREB/CREB and BDNF in the WT and KO groups. We found that the level of cAMP was significantly lower and the expression levels of pCREB/CREB and BDNF were significantly decreased in the KO group (Figures 5E,F). We also examined the cAMP/CREB/BDNF signaling pathway in FMT recipient mice and found that compared to the WT-FWT group, the level of cAMP was significantly decreased and the protein expression levels of pCREB/CREB and BDNF were significantly decreased in the WT-FKO group (Figures 5G,H). These results suggest that the cAMP/CREB/BDNF signaling pathway is inhibited in the KO group, leading to reduced expression of neurotrophic factors. Additionally, we examined the expression of three other common neurotrophic factors (CTNF, TGF-β, and NGF) in the KO and WT groups, and found that the expression of these factors showed a downward trend in the KO group compared to the WT group. However, after ABX treatment, the differential expression disappeared (Figure 5I). Similarly, WT-FKO mice receiving fecal bacteria from KO mice also showed significantly reduced expression of CTNF, TGF-β, and NGF (Figure 5J).
4. Discussion
Recent research has indicated that there is a close relationship between the balance of gut microbiota and depression. In this study, we found that Sig-1R knockout induced depression-like behaviors in mice, which was reversed upon clearance of gut microbiota with antibiotic treatment. Our findings were further supported by the observation that gut microbiota dysfunction and depression-like phenotypes in Sig-1R knockout mice could be reproduced through FMT experiments. Additionally, hippocampal RNA sequencing identified multiple KEGG pathways that are associated with depression. We also discovered that gut microbiota from Sig-1R-deficient mice may induce depression-like behaviors through regulation of the cAMP/CREB/BDNF signaling pathway. These results provide further evidence for the underlying mechanisms of depression and the link between the gut and the brain.
Sigma-1 receptor is highly expressed in brain regions related to mood. In our study, we found that Sig-1R knockout mice displayed a depression-like phenotype, including a significant reduction in immobility time and an increase in latency to immobility in the FST and TST, consistent with previous findings (Sabino et al., 2009). However, the mechanisms by which Sig-1R modulates antidepressant-like behaviors remain unclear. Previous studies have proposed that the serotonergic system may be involved (Szabo et al., 2021; Wu et al., 2021). Qin et al. (2022) found that inhibition of Sig-1R reduces PKC phosphorylation and suppresses GABAAR expression, leading to depression-like behaviors (Qin et al., 2022). Sha et al. (2015) also found that Sig-1R deficiency impairs neurogenesis, resulting in a depression-like phenotype (Sha et al., 2015). In our study, we found that the gut-brain axis may be a potential mechanism through which Sig-1R regulates depression-like behaviors. Sig-1R knockout mice displayed a depression-like phenotype that disappeared after depletion of gut microbiota with antibiotic treatment, and FMT could reproduce Sig-1R-mediated depression-like phenotypes. In recent years, neuroscientific research has demonstrated the importance of microbiota in depression (Evrensel and Ceylan, 2015; Chang et al., 2022). It has been reported that anxiety, depression, and aggressive behaviors can occur in mice with altered gut microbiota. FMT from various rodent models, such as unpredictable chronic mild stress (UCMS) models (Chevalier et al., 2020) and DSS models (Zhang F. et al., 2022), can reproduce depression-like phenotypes. This is the first study to confirm that feces from Sig-1R knockout mice had the same effect. Notably, no effect was observed on the SPI of the SPT after FMT treatment. We speculate that Sig-1R knockout did not cause severe depressive symptoms, which might be reflected in the SPT. Anhedonia in the sucrose preference test is defined as a preference for sucrose <65% (Scheggi et al., 2018; Fonseca-Rodrigues et al., 2022). Sig-1R knockout caused only minor variations in SPI, which may have been masked by FMT treatment.
Increasing evidence supports the connection between depression and the gut microbiota (Valles-Colomer et al., 2019; Yang et al., 2019). Depression is associated with decreased richness and diversity of the gut microbiota (Kelly et al., 2016). The pattern of the gut microbiome differs significantly between patients with depression and healthy controls (Barandouzi et al., 2020; Bertsch et al., 2022). In our study, Sig-1R knockout significantly altered the composition of the gut microbiota. At the genus level, the abundance of Alistipes, Alloprevotella, and Lleibacterium decreased significantly, and these changes were also observed in mice receiving FMT treatment. Alistipes, belonging to the family Rikenellaceae, has been shown to be significantly associated with depression (Jiang et al., 2015; Zhong et al., 2022). Alistipes has also been linked to depression-like phenotypes in other mood disorders (Domènech et al., 2022; Zhang et al., 2022a). In patients taking antidepressants, the abundance of Alistipes increased significantly, suggesting that it may play a crucial role in antidepressant effects (Chen Q. et al., 2021). Alloprevotella, which produces short-chain fatty acids and indole derivatives that may be involved in depression (Zheng et al., 2021), is more abundant in healthy patients compared to those with depression (Chen et al., 2022; Zhang K. et al., 2022). Depression-like symptoms in mice subjected to unpredictable chronic mild stress were relieved after CD36 knockout, and analysis of gut microbiota in CD36 KO mice revealed a significant increase in the abundance of Alloprevotella (Bai et al., 2021). A study on postpartum depression also found that Alloprevotella was associated with depressive phenotypes (Tian et al., 2021). A decrease in the relative abundance of Lleibacterium, which may be involved in enzymes that participate in purine metabolism, was observed in patients with major depression (Jiang et al., 2015). A reduced abundance of Lleibacterium was also found in our study. A recent study indicated that the abundance of Lleibacterium increased after consumption of dechlorogenic sunflower seeds and was negatively associated with depression-like phenotypes and markers of mucosal barrier damage (Lu et al., 2022).
ABX treatment has been demonstrated to alter gut microbiota composition and diversity dramatically in the host (Kennedy et al., 2018). To examine the role of gut microbiota in depression-like behaviors, ABX treatment was used to deplete gut microbiota. In our experiment, depression-like behaviors of Sig-1R KO mice were alleviated after ABX treatment, with changes in neurotrophic factors, suggesting that the disruption of gut microbiota caused by Sig-1R KO may play a significant role in the occurrence of depression-like behaviors. Although there was no statistically significant difference in immobility time in TST and latency to immobility in FST, it also showed a downward trend, which may be limited due to the restricted sample size. Consistent with our results, anhedonic-like phenotypes in chronic social defeat stress (CSDS) mice were alleviated after ABX treatment (Wang et al., 2020), suggesting that ABX-induced microbiome failure leads to stress resilience. Wang also found that antibiotic treatment alleviated depression-like behavior and decreased prefrontal cortical synaptic proteins in CSDS mice (Wang S. et al., 2021).
The vagus nerve system regulates the bidirectional communication between the gut microbiota and the brain. Extensive research has demonstrated that subdiaphragmatic vagotomy (SDV) can ameliorate depression-like behaviors. Subdiaphragmatic vagotomy block the depression-like phenotypes in ABX-treated mice with FMT from α7 subtype of the nicotinic acetylcholine receptor (Chrna7) KO mice (Pu et al., 2021). Behavior abnormalities in mice transplanted with CSDS fecal bacteria were significantly blocked by SDV (Wang et al., 2020). In our study, we found that the cAMP/CREB/BDNF signaling pathway was substantially downregulated in mice transplanted with Sig-1R KO fecal bacteria. However, the potential mechanism by which gut microbiota affects the brain remains unknown. The vagus nerve is associated with the development of neurological diseases by regulating immunity and inflammation. The association between the vagus nerve and depression-like behavior mediated by Sig-1R KO gut microbiota seems to be a research direction worth exploring.
There is a growing body of evidence linking BDNF to the pathogenesis of depression. In our study, lower levels of BDNF were also observed in Sig-1R knockout mice and mice transplanted with feces from Sig-1R knockout mice, compared to control mice. BDNF levels are significantly lower in patients with major depression than in healthy controls, and they recover after antidepressant treatment (Chen et al., 2001). This may be related to BDNF’s role in regulating synaptic plasticity and neurogenesis (Björkholm and Monteggia, 2016; Castrén and Kojima, 2017; Colucci-D’Amato et al., 2020). Additionally, we examined several other neurotrophic factors, including CTNF, TGF-β, and NGF, and found that their mRNA expression levels were also significantly reduced in Sig-1R knockout mice and FMT-KO mice. Decreased plasma levels of TGF-1 are associated with depression severity in patients with MDD (Caraci et al., 2018). Imbalances between IL-6 and TGF-β and between Th17 and Treg were also found in depressed patients or in animal models of depression, which induces neuroinflammation and neuronal dysfunction (Huang et al., 2022). These neurotrophic factors play crucial roles in memory formation and synaptic plasticity and have been identified as potential targets for the treatment of depression (Oglodek et al., 2016; Mondal and Fatima, 2019).
Brain-derived neurotrophic factor is a direct target of CREB (Barco et al., 2005), and cAMP-activated phosphorylated CREB regulates the expression of neurotrophic factors such as BDNF and NGF (Ortega-Martinez, 2015). The cAMP/CREB/BDNF pathway has been shown to be an important mechanism of action in traditional Chinese medicine (Cai et al., 2022) and novel antidepressants such as vortioxetine (Ramezany et al., 2019). In the hippocampal mRNA sequencing of Sig-1R knockout mice, the cAMP signaling pathway was significantly enriched. We also verified that the cAMP/CREB/BDNF signaling pathway was significantly downregulated at the protein level in Sig-1R knockout mice. FMT from Sig-1R knockout mice also resulted in downregulation of the cAMP/CREB/BDNF signaling pathway. This may be a potential mechanism by which the microbiome mediates depression-like behaviors. Through PPI network analysis of mRNA sequencing, we identified two core differentially expressed, Grin1 and Gria1. Genetic changes in Grin1 may increase the risk of depression (Weder et al., 2014). Nucleotide polymorphisms of Gria1 are associated with serotonin reuptake disorders in patients with depression (Bishop et al., 2012).
In our study, we found that the gut-brain axis may be a potential mechanism through which Sig-1R regulates depression-like behaviors. Our study is the first to provide evidence that fecal microbiota from Sig-1R knockout mice can induce depressive-like behavior in recipient mice. Our study offers new insights into the complex interplay between the gut and brain, specifically regarding the role of Sig-1R in regulating depressive-like behaviors.
5. Conclusion
In this study, we showed that Sig-1R knockout induced increased depression-like behaviors in mice with decreased neurotrophic factors, which may be mediated by gut microbiota. The microbiota from Sig-1R knockout mice was sufficient to cause a depression-like phenotype and a reduction in neurotrophic factors. The gut microbiota may regulate the expression of neurotrophic factors through the cAMP/CREB/BDNF signaling pathway (Figure 6). Our study offers a new perspective on the mechanism of action of Sig-1R in the regulation of depression and provides further evidence for the study of the gut-brain axis.
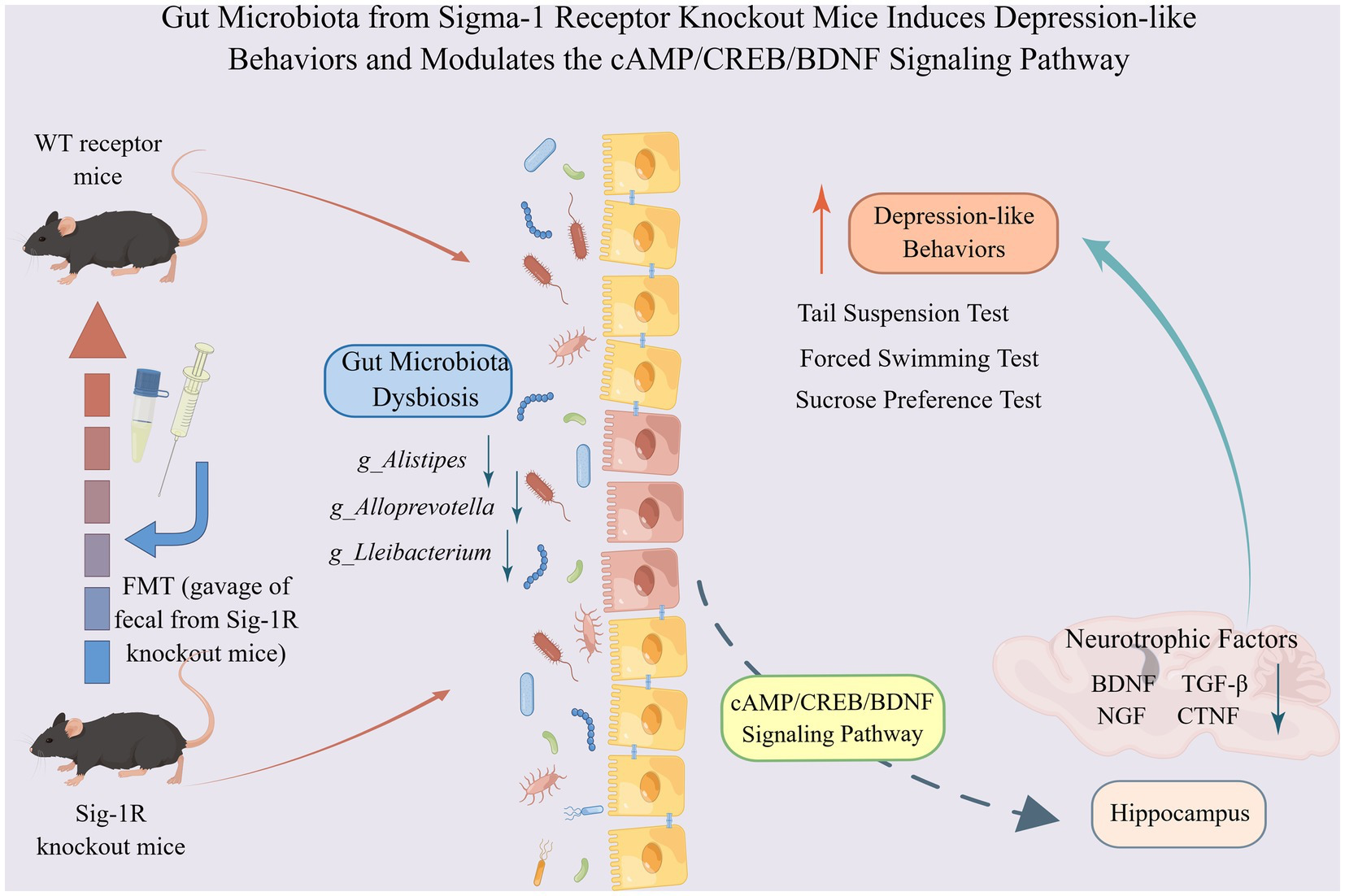
Gut microbiota from Sigma-1 receptor knockout mice induces depression-like behaviors and modulates the cAMP/CREB/BDNF signaling pathway. Sig-1R knockout induced increased depression-like behaviors in mice with decreased neurotrophic factors, which may be mediated by gut microbiota. The microbiota from Sig-1R knockout mice was sufficient to cause a depression-like phenotype and a reduction in neurotrophic factors. Gut microbiota may regulate the expressions of neurotrophic factors through the cAMP/CREB/BDNF signaling pathway. By Figdraw.