
Gut microbiota-derived short-chain fatty acids ameliorate methamphetamine-induced depression- and anxiety-like behaviors in a Sigmar-1 receptor-dependent manner
By Kaikai Zhang, Lijian Chen, Jianzheng Yang, Jiali Liu, Jiahao Li, Yi Liu, Xiuwen Li, Long Chen, Clare Hsu, Jiahao Zeng, Xiaoli Xie, and Qi Wanga
Excerpt from the article published in Acta Pharm Sin B. 2023 Dec;13(12):4801-4822. Epub 2023 Sep 21. PMID: 38045052; PMCID: PMC10692394. DOI: 10.1016/j.apsb.2023.09.010
Editor’s Highlights
- Methamphetamine (Meth) is a highly addictive psychostimulant.
- Meth-induced anxiety and depression relied on the inhibition of SIGMAR1/BDNF/TRKB pathway.
- Gut microbiota could contribute to the modulation of SIGMAR1 on behaviors.
- Gut microbiota participated in Meth-induced colonic inflammation and contributed to Meth-related anxiety and depression disorders.
- Activation of SIGMAR1 upregulated the BDNF/TRKB pathway and ameliorated Meth-induced depression- and anxiety-like behaviors.
- This further confirmed that Meth-induced mental disorders relied on its repression of SIGMAR1.
Abstract
Methamphetamine (Meth) abuse can cause serious mental disorders, including anxiety and depression. The gut microbiota is a crucial contributor to maintaining host mental health. Here, we aim to investigate if microbiota participate in Meth-induced mental disorders, and the potential mechanisms involved. Here, 15 mg/kg Meth resulted in anxiety- and depression-like behaviors of mice successfully and suppressed the Sigma-1 receptor (SIGMAR1)/BDNF/TRKB pathway in the hippocampus. Meanwhile, Meth impaired gut homeostasis by arousing the Toll-like receptor 4 (TLR4)-related colonic inflammation, disturbing the gut microbiome and reducing the microbiota-derived short-chain fatty acids (SCFAs). Moreover, fecal microbiota from Meth-administrated mice mediated the colonic inflammation and reproduced anxiety- and depression-like behaviors in recipients. Further, SCFAs supplementation optimized Meth-induced microbial dysbiosis, ameliorated colonic inflammation, and repressed anxiety- and depression-like behaviors. Finally, Sigmar1 knockout (Sigmar1−/−) repressed the BDNF/TRKB pathway and produced similar behavioral phenotypes with Meth exposure, and eliminated the anti-anxiety and -depression effects of SCFAs. The activation of SIGMAR1 with fluvoxamine attenuated Meth-induced anxiety- and depression-like behaviors. Our findings indicated that gut microbiota-derived SCFAs could optimize gut homeostasis, and ameliorate Meth-induced mental disorders in a SIGMAR1-dependent manner. This study confirms the crucial role of microbiota in Meth-related mental disorders and provides a potential preemptive therapy.
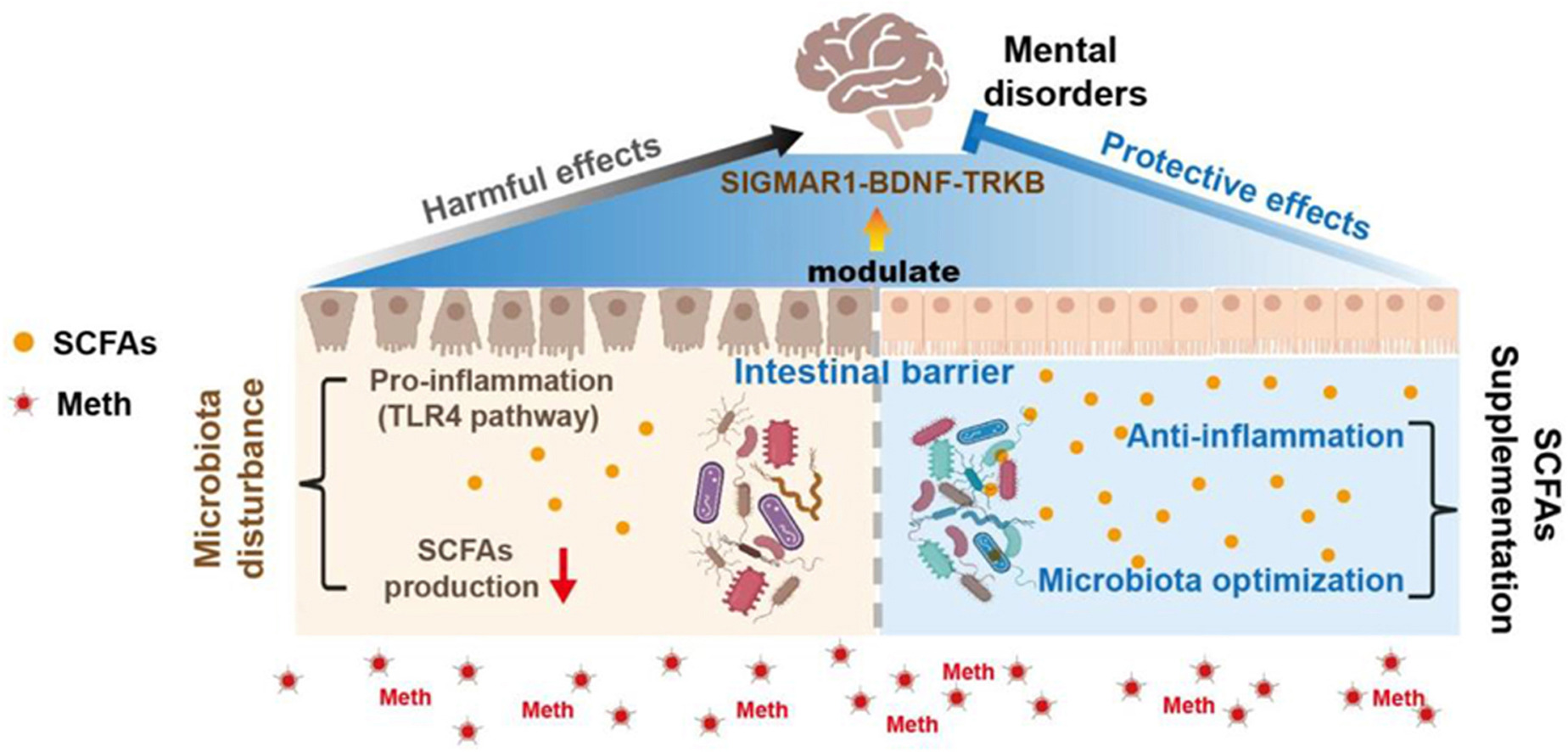
Graphical abstract
Gut microbiota involved in Meth-induced colonic inflammation and depression- and anxiety-like behaviors, while SCFAs optimized gut homeostasis and ameliorated mental disorders by modulating the SIGMAR1/BDNF/TRKB pathway.
1. Introduction
Methamphetamine (Meth) is a highly addictive psychostimulant, and its abuse has been a serious public health problem, globally1. According to an epidemiological survey, in 2019, there were over 27 million people reported using Meth (0.5% of the global population)2. Meth abuse can cause long-term neuropsychiatric impairments3, 4, 5, while anxiety and depression were reported as the most pervasive mental disorders in Meth abusers6,7.
Anxiety and depression are ubiquitous mental disorders that affect about 10% global population every year8. Accumulating evidence indicates the close association of gut microbiota with anxiety and depression disorders9, 10, 11. Clinically, microbial dysbiosisis prevalent in patients with depression and is recognized as a risk factor for disease12. Fecal microbiota transplantation (FMT) from depression patients confirmed its crucial role in the occurrence and development of depression10. In addition, the absence of microbiota could reduce the anxiety-like phenotypes13 and microbial metabolite 4-ethylphenyl sulfate has been confirmed to promote anxiety14. Notably, the interactions between gut microbiota and the brain are highly complex and involve multiple pathways, such as host metabolism, immune regulation, and vagal nerve pathway15, 16, 17, 18, 19. Through decomposing dietary fibers, microbiota can produce short-chain fatty acids, which maintain intestinal immune homeostasis20,21 and influence brain behaviors22,23. Aging-related decrease in SCFAs induced memory impairment and increased depression-like behavior24. Meanwhile, SCFAs presented a close association with chronic stress-induced anxiety25 and could attenuate chronic social failure stress-mediated depression26. These indicate that microbiota-derived SCFAs could be the potential modulator of multiple mental disorders.
Our previous studies have confirmed that Meth exposure can disturb gut microbiota, which participated in Meth-related hepatotoxicity2. In addition, in Meth-related conditioned place preference, fecal SCFAs were observed reshaped, accompanied by microbial dysbiosis27. Further, the absence of gut microbiota deteriorated the conditioned place preference scores28, implying the participation of microbiota in Meth-induced neurotoxicity. However, the microbial mechanisms underlying Meth-induced anxiety and depression, and the potential therapeutic effects of SCFAs remain largely unknown.
Sigma-1 receptor (SIGMAR1) is an endoplasmic reticulum chaperone protein widely expressed throughout the brain, which has shown protective effects against multiple mental disorders, including anxiety and depression29, 30, 31, 32. Its agonist fluvoxamine(Flu) has served as a clinical anti-anxiety and -depression drug33. In fact, the activation of SIGMAR1 can promote the secretion of mature brain derived neurotrophic factor (BDNF) from pro-BDNF via chaperone activity in the endoplasmic reticulum34, while BDNF and its receptor tyrosine kinase B (TRKB) were the crucial anti-anxiety and -depression factors35. The beneficial effects of SIGMAR1 on Meth-induced neural deficits were also observed, including ameliorating Meth-induced dopamine efflux, repressing drug-seeking brain reward function, and improving locomotor sensitization36. Nevertheless, the protective effects of SIGMAR1 against Meth-related anxiety and depression disorders, and its interaction with microbiota-derived SCFAs are still obscure.
In this study, we tested the hypothesis that microbial dysbiosis contributes to Meth-related anxiety and depression, while microbiota-derived SCFAs could modulate these mental disorders in SIGMAR1 dependent manner. Through FMT treatment, we confirmed the involvement of microbial dysbiosis in Meth-induced colonic inflammation, and anxiety and depression. By SCFAs supplementation, we verified its optimization on gut homeostasis and its protection against Meth related-anxiety and -depression. Finally, using Sigmar1−/− mice and agonist Flu, we found that the effects of these interventions on Meth-induced mental disorders relied on the modulation of the SIGMAR1/BDNF/TRKB pathway.
…
3. Results
3.1. Meth induced anxiety- and depression-like behaviors in mice and repressed the SIGMAR1/BDNF/TRKB pathway of the hippocampus
Meth exposure can cause anxiety and depression in abusers. Here, mice were administrated 15 mg/kg Meth to induce anxiety- and depression-like behaviors (Fig. 1A). We observed that compared to the Saline group, Meth significantly lowered the body weight of mice (Fig. 1B). In the OFT, Meth administration had a significant inhibitory effect on the total travel distance and the time spent in the center zone and displayed an increase in fecal number in comparison with the Saline mice (Fig. 1C and D; Supporting Information Fig. S1A). Meanwhile, the EPMT results showed that the time mice spent in and the number of entries into the open arms was reduced after Meth administration (Fig. 1C and D; Fig. S1B). In addition, Meth significantly elevated immobility time during the TST and FST relative to the Saline group (Fig. 1E). These results indicated that this dosing regimen successfully resulted in anxiety- and depression-like behaviors in mice.

Meth induced anxiety- and depression-like behaviors of mice, and inhibited the SIGMAR1/BDNF/TRKB pathway in the hippocampus.
(A) Schematic diagram of the experimental procedure. (B) Body weight change curves from three days prior to dose initiation till the end of final dose administration (n = 9–10 per group). (C) The representative trajectory diagrams in the OFT and EPMT. (D) The time mice spent in the center zone during the OFT and the time mice spent in the open arms during the EPMT (n = 10 per group). (E) The total immobility time during the TST and FST (n = 11–12 per group). (F) IF staining of SIGMAR1 (in green), BDNF (in green), and p-TRKB (in red). Nuclei were labeled with DAPI (in blue). The number of positive cells was shown (n = 6 per group). Scale bar, 50 μm. (G) Expression of proteins SIGMAR1, BDNF, p-TRKB, the total TRKB, and β-actin was analyzed by Western blot. β-Actin served as a loading control. n = 3 per group. Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
Given the protective effects of SIGMAR1 on anxiety and depression, we determined the expression of the SIGMAR1/BDNF/TRKB pathway in the dentate gyrus (DG) region of the hippocampus, which crucially contributed to anxiety and depression regulation53. IF analysis showed that Meth reduced the number of positive cells SIGMAR1, BDNF, and p-TRKB (Fig. 1F). Western blot analysis confirmed these alterations: the expression of proteins SIGMAR1, BDNF, and p-TRKB was reduced after Meth administration in comparison with the Saline mice, and the level of the total TRKB was not altered (Fig. 1G). At the mRNA level, Meth also lowered the level of Sigmar1 and Bdnf, while it had no influence on the expression of mRNA Trkb (Fig. S1C), which was consistent with the protein expression of the total TRKB. Thus, Meth exposure could inhibit the SIGMAR1/BDNF/TRKB pathway in the hippocampus. In addition, we also determined the expression of SIGMAR1 in some other brain regions, including the prefrontal cortex, striatum, and midbrain. Both IF and Western blot analyses revealed no significant difference in SIGMAR1 protein expression in the prefrontal cortex and striatum after Meth exposure compared to the saline group. However, a decrease was observed in the midbrain (Fig. S1D–S1I), suggesting region-specific effects of Meth on SIGMAR1 expression in the brain.
3.2. Meth impaired the gut homeostasis by arousing colonic inflammation, disturbing gut microbiota, and reshaping the composition of SCFAs
The microbiota–gut–brain axis has emerged a crucial role in the regulation of multiple mental disorders, including anxiety and depression9, 10, 11. Thus, we determined the impacts of Meth on gut homeostasis.
3.2.1. Meth damaged the intestinal barrier function and aroused TLR4-related colonic inflammation
As shown in Fig. 2A, Meth significantly reduced the length of the colon relative to the Saline group. HE staining displayed that Meth caused the larger volume and the “balloon-like” changes of goblet cells (Fig. 2B). Further, AB and PAS staining presented an increased number of AB+ and PAS+ goblet cells after Meth administration (Fig. 2B), indicating that Meth promoted the hyperplasia of goblet cells and increased mucus secretion. Meanwhile, we observed damaged intestinal barrier function after the Meth group, as demonstrated by increased intestinal permeability to FITC-dextran and serum LPS level (Fig. 2C).

Meth aroused colonic inflammation by activating the TLR4/NF-κB pathway.
(A) The representative images and the length of the colon in the Saline and Meth groups were shown (n = 5 per group). (B) The representative images of HE, AB, and PAS staining of colonic sections from mice. The number of AB+ and PAS+ goblet cells per crypt was shown (three random visual fields for each sample, n = 15 per group). Scale bar, 150 μm. (C) The concentration of FITC-dextran and LPS in serum (n = 5–7 per group). (D) PCA plot analysis of Saline (green dots) and Meth samples (red squares) (n = 3 per group). (E) The Scatter-plot graphs of the differential expression genes between Saline and Meth groups: the red rhombuses represented upregulated genes, the green squares represented downregulated genes and the grey dots represented no significant difference. (F) GO analysis of DEGs in the colon. DEGs were annotated to 10 biological processes (red), 7 cellular components (blue), and 3 molecular functions (green) according to the functional classification. (G) KEGG enrichment analysis of DEGs. Red font represents the pathways belonging to the second category of the Immune system; orange font belongs to the Infectious disease: bacterial; wathet blue font belongs to the Infectious disease: viral; green font belongs to the Infectious disease: parasitic; dark blue font belongs to the Cell growth and death; purple font belongs to the Nervous system; black font belongs to the Substance dependence. (H) Heatmaps of gene expression profiles enriched into the Toll-like receptor signaling pathway and the NOD-like receptor signaling pathway. (I) PPI analysis of DEGs enriched into these two pathways. (J) Expression of proteins TLR4, p-NF-κB, the total NF-κB, ZO-1, OCCLUDIN, and β-actin was analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). (K) Relative expression of mRNA Tnf-α, Il-6, and Il-1β was normalized by β-actin mRNA (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
o further reveal the molecular mechanisms of Meth-induced colonic impairment, mRNA sequencing was performed. According to the principal component analysis, Meth exposure led to the alteration of colonic components relative to the Saline group (Fig. 2D). Through the screening of DEGs, Meth resulted in a total of 1246 DEGs in the colon relative to the Saline group, containing 693 up-regulated and 553 down-regulated DEGs (Fig. 2E). Furthermore, 87 pathways involved in Meth-induced colonic toxicity were enriched based on KEGG analysis (Supporting Information Table S3). The immune system was identified both in KEGG and GO annotation analysis (Fig. 2F and G; Supporting Information Fig. S2), while the inflammation-related Toll-like receptor signaling pathway and NOD-like receptor signaling pathway were also enriched (Fig. 2H), implying the occurrence of colonic inflammation. Interestingly, Amphetamine addiction was significantly enriched, implying the involvement of the gut–brain axis in Meth addiction(Fig. 2G). Additionally, Tlr4 was identified as the core gene by the PPI analysis (Fig. 2I), which was the classical target of microbiota-derived LPS54. Correspondingly, the expression of protein TLR4 and its downstream nuclear phospho-nuclear factor-κB (p-NF-κB) was upregulated by Meth, while the total NF-κB remained unchanged (Fig. 2J). Meanwhile, Meth elevated the expression of inflammatory factors at the mRNA level, including tumor necrosis factor-α (Tnf-α), interleukin-6 (Il-6), and interleukin-1β (Il-1β) (Fig. 2K). Corresponding to increased intestinal permeability, the tight junction proteins zona occludens 1 (ZO-1) and OCCLUDIN were significantly down-regulated after Meth administration (P-value < 0.05) (Fig. 2J). These results indicated that Meth could arouse colonic inflammation by activating the TLR4/NF-κB pathway and damage the intestinal barrier function.
3.2.2. Meth disturbed gut microbiota and reshaped the composition of SCFAs
16S rRNA sequencing was performed to profile the gut microbiome. Rank-Abundance curves presented similar OTU abundance or evenness among all samples (Fig. 3A). In Alpha diversity, Meth exposure resulted in a higher richness than the Saline group (up-regulated ACE, Chao, and Sobs indexes), while no changes were observed in microbial diversity and coverage (demonstrated by Shannon, Simpson, and Coverage indexes) (Fig. 3A; Supporting Information Fig. S3A–S3E). In Beta diversity, the principal composition of Meth samples was significantly different from that of the Saline group (Fig. 3B), which was in accord with the Hierarchical clustering analysis (Fig. S3F). At the Phylum level, Meth induced a higher abundance of Firmicutes and Deferribacterota, as well as the lower Bacteroidate and Proteobacteria (Fig. 3C; Fig. S3G). Moreover, the ratio of Firmicutes/Bacteroidate was significantly upregulated by Meth administration (Fig. 3D), indicating a worse intestinal environment55. At the Family level, Meth altered the abundance of 17 microbes, including 10 upregulated (major Lactobacillaceae, Rikenellaceae, and Deferribacteraceae) and 7 down-regulated microbes (major Muribaculaceae, Sutterellaceae, and Erysipelotrichaceae) (Fig. S3H–S3I). At the Genus level, the SCFAs producers Lactobacillus, Lachnospiraceae_NK4A136_group, and pathogenic Alistipes and Mucispirillum were increased by Meth administration, whereas probiotics norank_f_Muribaculaceae and Faecalibaculum were decreased (Fig. 3E; Fig. S3J). The differential microbiota from Phylum to Genus level were summarized using the LEfSe cladogram and linear discriminant analysis (Fig. 3F and G). The Sankey diagram further revealed the relative abundance of differential microbiota at Phylum and their taxon relationship with the differential microbiota at the Genus level (Fig. S3K). These results suggested that Meth could disturb the gut microbiome and alter SCFAs-related microbes.

Meth disturbed the gut microbiome and reshaped the composition of fecal SCFAs.
(A) The Rank-Abundance Curves of all samples: the abscissa denoted the ranking level of OTU number and the corresponding ordinate denoted the relative percentage; and the alteration of alpha diversity: ACE index at OTU level (n = 8–9 per group). (B) PCoA plot analysis. (C) Relative abundance of microbiota at the Phylum level. (D) The ratio of phylum Firmicutes/Bacteroidate. (E) The top 15 differential microbiota between the Saline and Meth groups at the genus level. (F) LEfSe cladogram indicated that multiple taxa are differentially enriched in the corresponding groups. The circles from the inner to the outer circles indicate the phylum, class, order, family, and genus levels. (G) Histogram of linear discriminant analysis (LDA) in the abundance of gut microbiota. Wilcoxon rank-sum test with a 95% confidence interval was performed to screen the differential microbiota. (H) The concentration of SCFAs in the Saline and Meth group, including acetic acid, propanoic acid, butanoic acid, valeric acid, hexanoic acid, isobutyric acid, isohexanoic acid, and isovaleric acid (n = 6–8 per group). (I) Spearman correlation analysis of differential microbiota with colonic, behavioral indicators, SCFAs concentration, and IF. The color of the dots denoted the correlation coefficient and the size of the dots represented the P-value. Red dots represent a positive correlation, while the blue ones represent a negative correlation. Data are expressed as mean ± SEM, ∗P <0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05).
Thus, we determined the composition of SCFAs in feces. Among 8 SCFAs, Meth effectively decreased the level of acetic acid, propanoic acid, valeric acid, isobutyric acid, and isovaleric acid, while there were no significant changes in butanoic acid, hexanoic acid, and isohexanoic acid (Fig. 3H). Spearman correlation analysis was performed to reveal the potential association between gut microbiota (Genus level) and SCFAs concentration, behavioral indexes, colonic indexes, and IF indexes (Fig. 3I). The results showed that the abundance of norank_f_Muribaculaceae, Faecalibaculum, norank_f_Desulfovibrionaceae, Helicobacter, Alloprevotella, Dubosiella, Bifidobacterium, Parasutterella, and Desulfovibrio was positively correlated with body weight, colonic length, fecal SCFAs concentration and the expression of SIGMAR1, BDNF, and p-TRKB, while negatively correlated with the number of AB+ and PAS+ goblet cells, FD4 level and the depression- and anxiety-like behaviors. Meanwhile, the abundance of Lactobacillus, Alistipes, Lachnospiraceae_NK4A136_group, Mucispirillum, Rikenellaceae_RC9_gut_group, and Roseburia presented a positive correlation with the FD4 level, the number of AB+ and PAS+ goblet cells, and the depression- and anxiety-like behaviors, while presented a negative correlation with the body weight, colonic length, SCFAs concentration and the expression of SIGMAR1, BDNF, and p-TRKB (Fig. 3I). These results indicated the potential modulation of gut microbiota on colonic damage and behavioral deficits. Furthermore, the composition of serum SCFAs was also assessed following Meth exposure. As observed in fecal samples, Meth primarily reduced the concentration of serum Acetic acid and propanoic acid (Supporting Information Fig. S4A and S4B), and did not exert a significant effect on butanoic acid (Fig. S4C). Additionally, the level of hexanoic acid in serum decreased with Meth administration, whereas there were no statistically significant differences in the levels of valeric acid, isobutyric acid, isovaleric acid, or isohexanoic acid (Fig. S4D–S4H).
3.3. Microbiota from Meth-administrated mice aroused colonic inflammation and further induced anxiety- and depression-like behavior in the recipient mice
Here, we transplanted the microbiota from Meth- and Saline-administrated mice into recipient mice to explore the underlying microbial mechanisms (Fig. 4A). Previously, we did not detect the concentration of Meth and its main metabolite-amphetamine in fecal samples, while we did detect them in serum samples (Meth: 3.728 ± 0.3496 ng/mL; amphetamine: 0.2737 ± 0.04802 ng/mL, mean ± SEM, n = 8)2. The results excluded the influences of drug residue in feces.
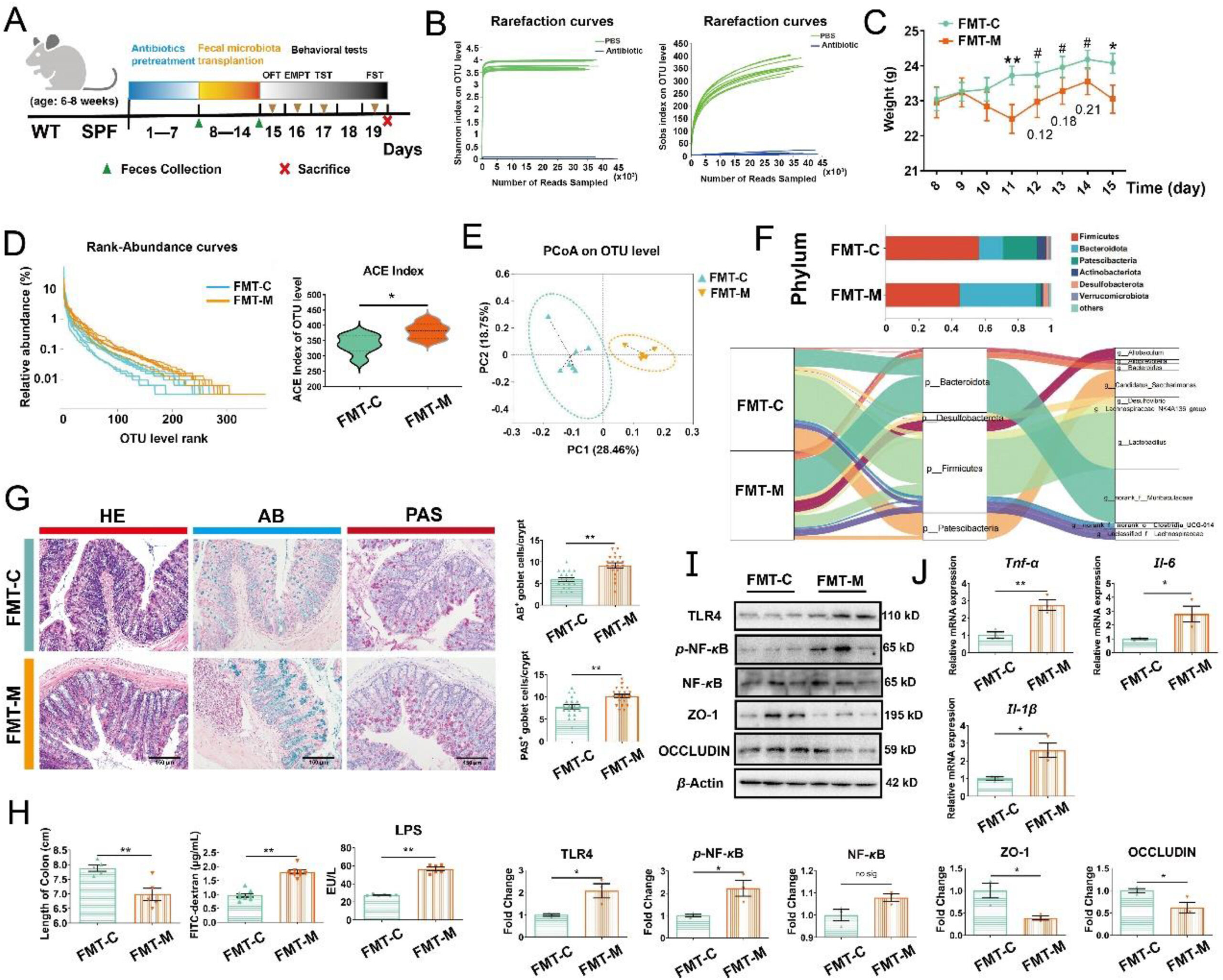
Microbiota from Meth-administrated mice induced colonic inflammation by activating the TLR4/NF-κB pathway.
(A) Schematic diagram of the experimental procedure. (B) Rarefaction curves of samples in PBS and Antibiotic group (n = 8 per group). (C) Body weight change curves (n = 7–9 per group). (D)The Rank-Abundance Curves of all samples and the alteration of alpha diversity: ACE index at the OTU level (n = 6–7 per group). (E) PCoA plot analysis. (F) Relative abundance of microbiota at the Phylum level and Sankey diagram: relative abundance of differential microbiota at the Phylum level and the corresponding flows of Genus differential microbiota. The color of the ribbon represented the differential microbiota and its width represented the relative abundance of differential microbiota. (G) The representative images of HE, AB, and PAS staining of colonic sections from mice (n = 6 per group). Scale bar, 150 μm. The number of AB+ and PAS+ goblet cells per crypt was shown (three random visual fields for each sample) (n = 20 per group). (H) The length of the colon (n = 5 per group), the concentration of FITC-dextran (n = 8 per group), and LPS in the serum (n = 6 per group). (I) Expression of proteins TLR4, p-NF-κB, total NF-κB, ZO-1, OCCLUDIN, and β-actin analyzed by Western blot (n = 3 per group). (J) Relative expression of mRNA Tnf-α, Il-6and Il-1β (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, #P >0.05, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
3.3.1. Microbiota from Meth-administrated mice aroused colonic inflammation and damaged intestinal barrier function
As the sequencing depth increased, the rarefaction curves of Shannon and Sobs indexes in antibiotic-pretreated mice were not elevated in comparison to the PBS-pretreated mice, suggesting the elimination of gut microbiota (Fig. 4B). Next, FMT was performed on the recipients. Compared with FMT-C, we observed that FMT-M lowered the body-weight of recipients from day 4 of FMT and this tendency continued until the end of the procedure (Fig. 4C). Corresponding to Meth-administrated mice, there was no significant difference in OTU abundance or evenness among samples (Fig. 4D). FMT-M also increased the richness of the microbiome by elevating the ACE, Chao, and Sobs indexes in comparison with the FMT-C group (Fig. 4D; Supporting Information Fig. S5A and S5B). And diversity was observed to increase by FMT-M: up-regulated Shannon index and down-regulated Simpson index (Fig. S5C and S5D). In addition, Beta diversity presented that the principal composition of FMT-M samples differed from FMT-C according to the PCoA analysis (Fig. 4E). The microbial composition of FMT-M from Phylum to Genus level was significantly different from FMT-C (Fig. 4F; Fig. S5E–S5G). The Sankey diagram further revealed the relative abundance of differential microbiota at the Phylum level and their taxon relationships with differential microbiota at the Genus level (Fig. 4F). These results indicated that we successfully transplanted the microbial community into the recipients.
Furthermore, FMT-M enlarged the size of goblet cells compared to FMT-C. The number of AB+ and PAS+ goblet cells was elevated by FMT-M (Fig. 4G), suggesting the promotion of proliferation in goblet cells. Moreover, FMT-M significantly reduced the length of the colon and increased the intestinal permeability, as demonstrated by the higher FITC-dextran and LPS level in serum (Fig. 4H). As observed in Meth-administrated mice, FMT-M also aroused colonic inflammation by up-regulating the expression of proteins TLR4 and nuclear p-NF-κB (the total NF-κB was not changed), as well as increasing the mRNA level of Tnf-α, Il-6, and Il-1β (Fig. 4I and J). Consisting of the increased intestinal permeability, the expression of ZO-1 and OCCLUDIN was decreased by FMT-M (Fig. 4I). Thus, we certified that gut microbiota participated in Meth-induced colonic inflammation via activating TLR4/NF-κB pathway and damaged intestinal barrier.
3.3.2. Microbiota from Meth-administrated mice repressed the SIGMAR1/BDNF/TRKB pathway and induced anxiety- and depression-like behaviors
Given the regulatory effects of microbiota on mental disorders56, we evaluated the extent of depression and anxiety after FMT-M administration. Behavioral results showed that FMT-M resulted in shorter total travel distance, lesser time spent in the center zone and more feces in OFT (Fig. 5A and B; Supporting Information Fig. S6A and S6B), lesser time spent in and fewer entries into the open arms in the EPMT (Fig. 5A and B; Fig. S6C) and longer immobility time of mice during the TST and FST compared with FMT-C mice (Fig. 5C). Western blot and IF analysis showed that FMT-M induced a lower expression of SIGMAR1, BDNF, and p-TRKB than those in the FMT-C group (Fig. 5D and E), while the total TRKB was not altered (Fig. 5D). Accordingly, Meth-induced microbial dysbiosis was involved in Meth-related depression and anxiety, while the repression of the SIGMAR1/BDNF/TRKB pathway could be a potential mechanism.
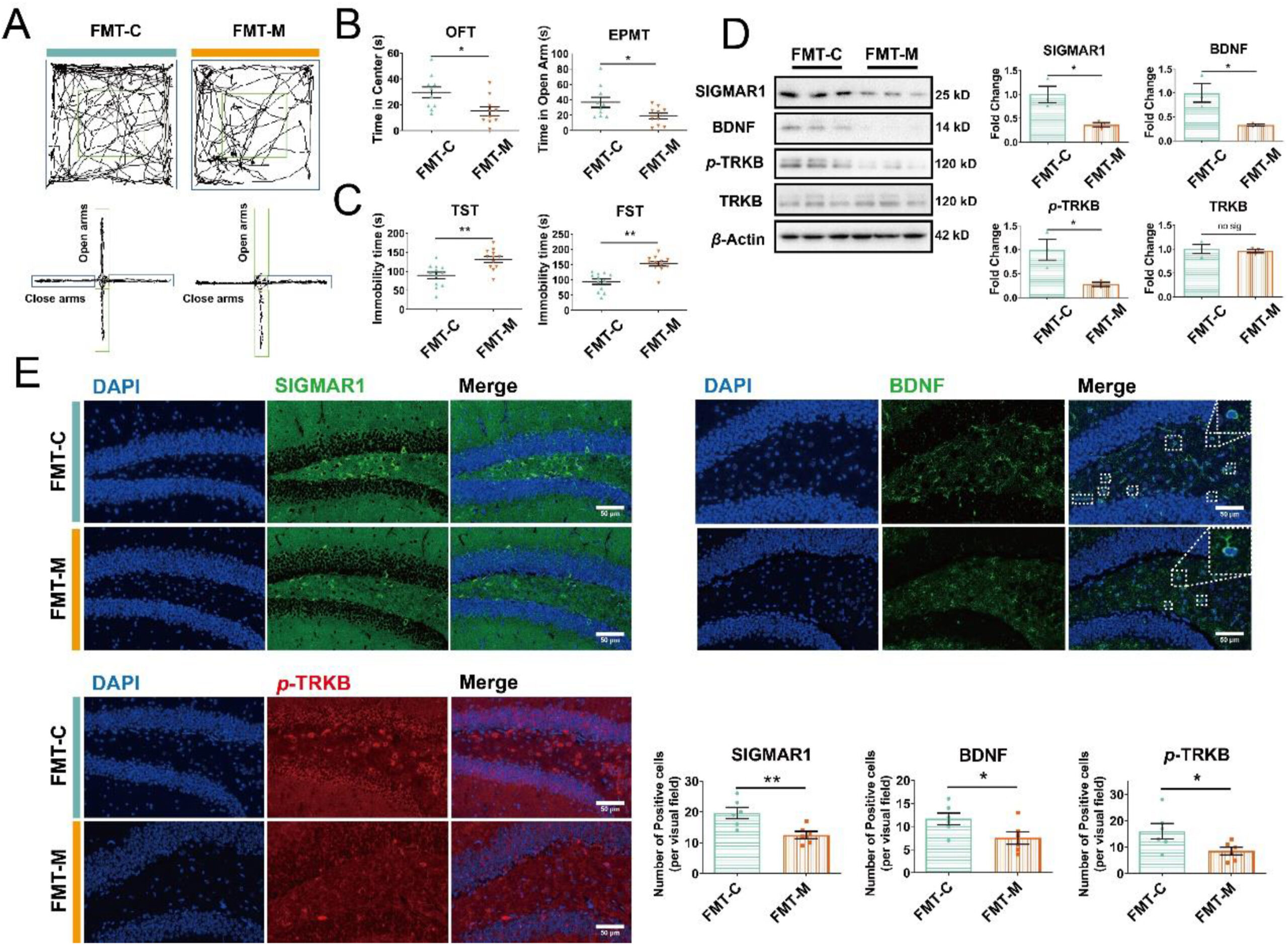
Figure 5.
Microbiota from Meth-administrated mice inhibited the SIGMAR1/BDNF/TRKB pathway in the hippocampus and induced anxiety- and depression-like behaviors of recipients.
(A) The representative trajectory diagrams in OFT and EPMT. (B) The time the mice spent in the center zone (OFT) and the time the mice spent in open arms (EPMT) (n = 10 per group). (C) The total immobility time during the TST and FST (n = 12 per group). (D) Protein expressions of SIGMAR1, BDNF, p-TRKB, the total TRKB, and β-actin in FMT-C and FMT-M samples were analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). (E) IF staining of SIGMAR1 (in green), BDNF (in green), and p-TRKB (in red) in FMT-C and FMT-M samples. Nuclei were labeled with DAPI (in blue). The number of positive cells is shown (n = 6 per group). Scale bar, 50 μm. Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
3.4. Microbiota-derived SCFAs attenuated Meth-induced colonic inflammation, and ameliorated the anxiety- and depression-like behaviors
Microbiota-derived SCFAs have emerged the beneficial effects on maintaining intestinal homeostasis and mental disorders20, 21, 22, 23. Here, acetic and propanoic acids were observed to decrease by Meth. Thus, we added the mixture of acetic and propanoic acids into the drinking-water of mice to investigate its potential protection (Fig. 6A).
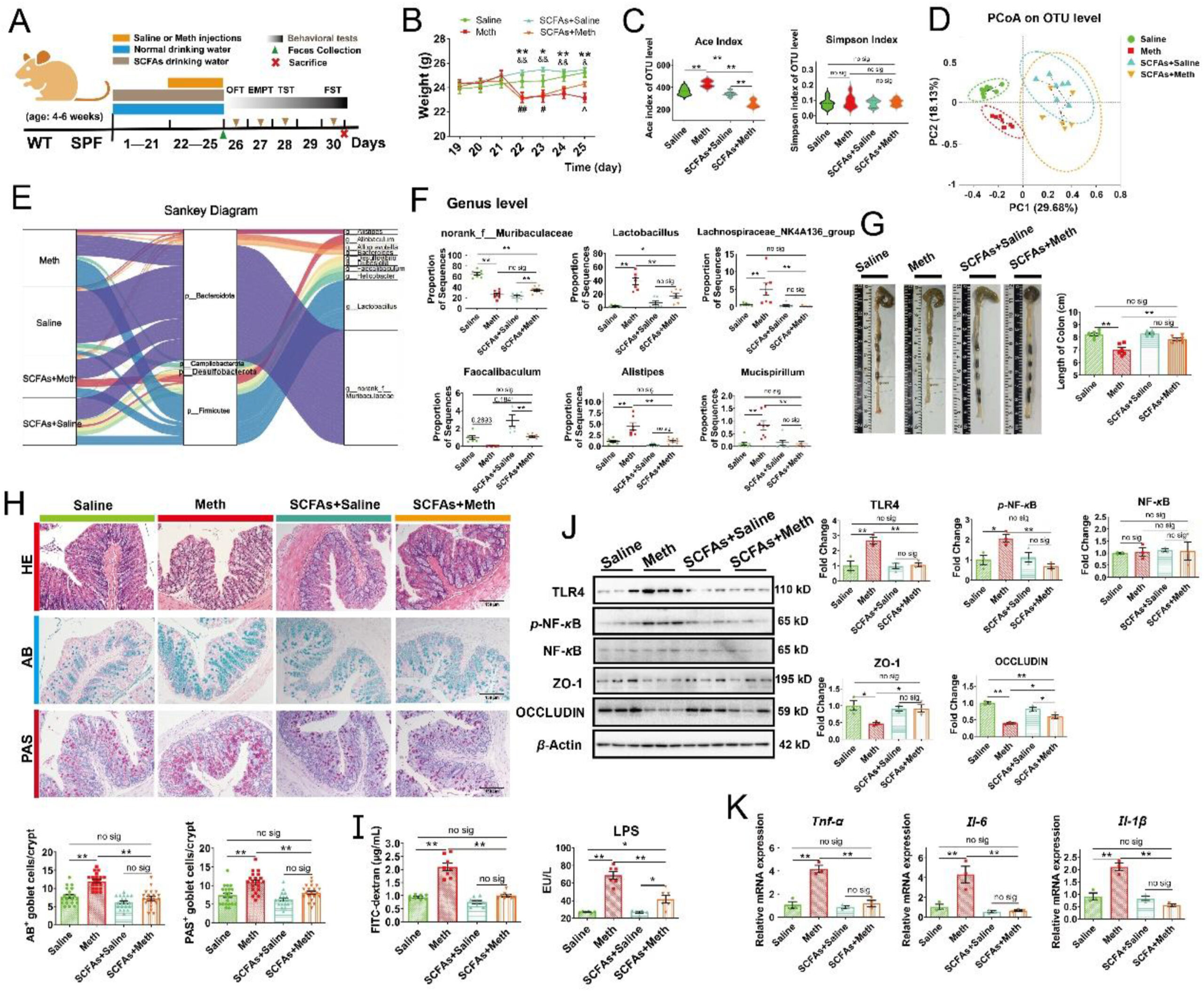
Microbiota-derived SCFAs optimized gut microbiota and protected against Meth-induced colonic inflammation.
(A) Schematic diagram of the experimental procedure. (B) Body weight change curves. ∗P < 0.05, ∗∗P < 0.01 (Meth vs. Saline group); #P < 0.05, ##P < 0.01 (SCFA + Meth vs. Saline group); &P < 0.05, &&P < 0.01 (SCFA + Meth vs.SCFA + Saline group); ˆP < 0.05, (SCFA + Meth vs. Meth group), (n = 6 per group). (C) The alteration of alpha diversity at the OTU level: ACE and Simpson indexes (n = 6–9 per group). (D) PCoA plot analysis at the OTU level. (E) Sankey diagram: relative abundance of differential microbiota at the Phylum level and the corresponding flows of Genus differential microbiota. The color of the ribbon represented the differential microbiota and its width represented the relative abundance of differential microbiota. (F) The proportion of microbiota sequences at the Genus level across each group. Kruskal–Wallis test with a 95% confidence interval was performed to screen the differential microbiota. (G) The representative images and the colon length across each group (n = 6 per group). (H) The representative images of HE, AB, and PAS staining of colonic sections from mice (n = 6 per group). The number of AB+ and PAS+ goblet cells per crypt across each group was shown (three random visual fields for each sample) (n = 20 per group). Scale bar, 150 μm. (I) The concentration of FITC-dextran (n = 8 per group) and LPS (n = 6 per group) in serum. (J) Protein expressions of TLR4, p-NF-κB, total NF-κB, ZO-1, OCCLUDIN, and β-actin across each group analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). (K) Relative expression of mRNA Tnf-α, Il-6, and Il-1β across each group was normalized by β-actin mRNA (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
3.4.1. Microbiota-derived SCFAs optimized the composition of the gut microbiome and protected against Meth-induced colonic inflammation
Meth initially caused significant weight loss in mice with or without SCFAs supplementation, while SCFAs accelerated its recovery (Fig. 6B). 16S rRNA sequencing revealed the similar OTU abundance or evenness among samples (Supporting Information Fig. S7A). In Alpha diversity, compared to the Saline group, Meth increased the Ace, Chao, and Sobs indexes, while SCFAs supplementation attenuated these alterations (Fig. 6C; Fig. S7B and S7C); meanwhile, there was no significant difference of Simpson and Shannon indexes across groups (Fig. 6C; Fig. S7D), indicating that SCFA could ameliorate Meth-induced increase of microbial richness, and presented no influences on diversity. In Beta diversity, SCFAs dominated the alteration of the principal component in fecal samples, which were further reshaped by Meth administration (Fig. 6D). Furthermore, the abundance of gut microbiota was changed by SCFAs supplementation from Phylum to Genus level (Fig. S7E–S7H). The Sankey diagram further revealed the relative abundance of differential microbiota at the Phylum level and their taxon relationships with differential microbiota at the Genus level in each group (Fig. 6E). At the Genus level, Meth significantly reduced the abundance of norank_f_Muribaculaceae, while SCFAs did not rescue this alteration (Fig. 6F). Meanwhile, SCFAs supplementation ameliorated the Meth-induced increase of Lactobacillus and normalized the abundance of Lachnospiraceae_NK4A136_group, pathogenic Alistipes and Mucispirillum and probiotics Faecalibaculum (Fig. 6F). These results suggested that SCFAs could reshape the gut microbiome and partly normalize Meth-induced microbial disturbance.
Additionally, we observed that SCFAs supplementation prevented Meth-induced shortening of colonic length (Fig. 6G). Correspondingly, SCFAs effectively alleviated Meth-induced enlargement and “balloon-like” changes of goblet cells (Fig. 6H). Further, Meth-induced increase in the number of AB+ and PAS+ goblet cells was also normalized by SCFAs supplementation (Fig. 6H), suggesting that SCFAs could repress Meth-induced excessive proliferation of goblet cells. Additionally, SCFAs presented protective effects on intestinal permeability: Meth induced the elevation of serum FITC-dextran and LPS levels, which were attenuated by SCFAs supplementation (Fig. 6I). In addition, SCFAs effectively attenuated Meth-induced colonic inflammation by inhibiting the activation of the TLR4/NF-κB pathway, evidenced by the lower level of proteins TLR4 and nuclear p-NF-κB, along with the lower Tnf-α, Il-6, and Il-1β at mRNA level than Meth samples (Fig. 6J and K). Meanwhile, the upregulation of proteins ZO-1 and OCCLUDIN were also observed by SCFAs supplementation relative to the Meth group (Fig. 6J). Thus, our results demonstrated that SCFAs could protect against Meth-induced colonic inflammation and improve intestinal barrier function.
3.4.2. Microbiota-derived SCFAs upregulated the SIGMAR1/BDNF/TRKB pathway and ameliorated Meth-induced anxiety- and depression-like behaviors of mice
Given the beneficial effects of SCFAs on multiple mental disorders, we also explored the regulation of SCFAs on Meth-induced anxiety- and depression disorders. Our results showed that SCFAs effectively mitigated the Meth-induced decrease in depression- and anxiety-like behaviors, demonstrated by the longer total travel distance, more time the mice spent in the center zone, and the decrease of feces amount in OFT (Fig. 7A and B; Supporting Information Fig. S8A and S8B); the more time mice spent in and the entries into open arms in EPMT (Fig. 7A and B; Fig. S8C); the lesser immobility time in TST and FST than Meth group (Fig. 7C). Moreover, the expression level of SIGMAR1, BDNF, and p-TRKB in Meth-administrated mice was significantly lower than in Saline mice, while SCFA supplementation could alleviate this downregulation (the total TRKB was not altered) (Fig. 7D–F), indicating that SCFAs could rescue Meth-induced inhibition of the SIGMAR1/BDNF/TRKB pathway. Altogether, these results indicated that SCFAs could effectively prevent Meth-induced depression- and anxiety-like behaviors by regulating the SIGMAR1/BDNF/TRKB pathway. Spearman correlation analysis revealed the potential regulation of the microbiota on the colonic damage, IF indexes, and depression- and anxiety-related indicators (Fig. 7G). Specially, the abundance of Faecalibaculum, norank_f_Desulfovibrionaceae, Dubosiella, and Desulfovibrio was strongly positively correlated with the body weight, colonic length, and the expression of SIGMAR1, BDNF, and p-TRKB, while negatively correlated with the number of AB+ and PAS+ goblet cells, FD4 level, and the depression- and anxiety-like behaviors. Meanwhile, the abundance of Lactobacillus, Alistipes, Rikenellaceae_RC9_gut_group, Bacteroides, and Roseburia presented a positive correlation with the FD4 level, the number of AB+ and PAS+ goblet cells, and the depression- and anxiety-like behaviors, while negatively correlated with the body weight, colonic length, and the expression of SIGMAR1, BDNF, and p-TRKB (Fig. 7G). These suggested that SCFAs-mediated optimization of gut microbiota could participate in its ameliorative effects on Meth-induced colonic damage and behavioral deficits.
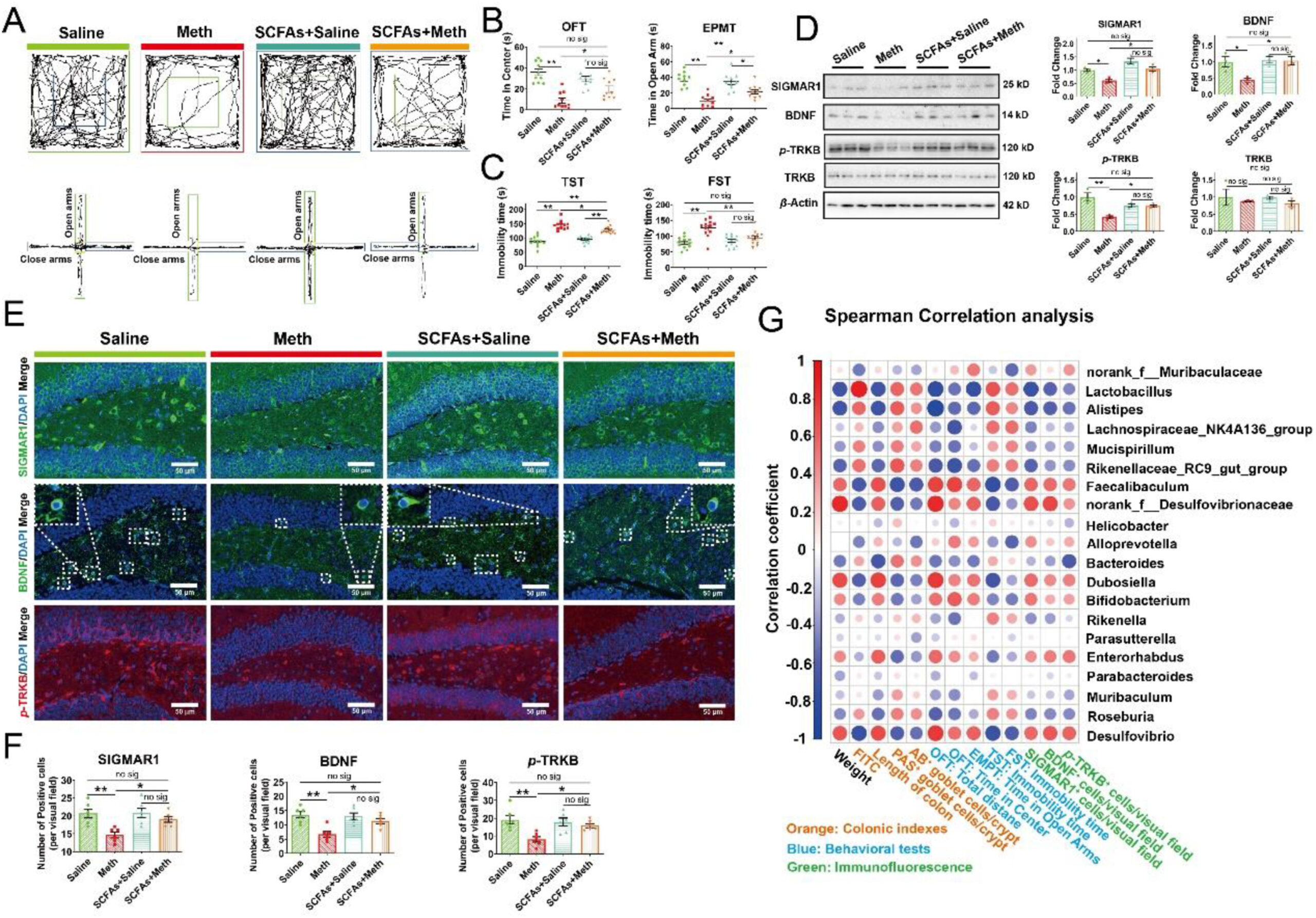
Microbiota-derived SCFAs upregulated the SIGMAR1/BDNF/TRKB pathway in the hippocampus and ameliorated Meth-induced anxiety- and depression-like behaviors.
(A) The representative trajectory diagrams in OFT and EPMT. (B) The time the mice spent in the center zone (OFT) and the time the mice spent in the open arms (EPMT) across each group (n = 10 per group). (C) The total immobility time across each group during the TST and FST (n = 12 per group). (D) Protein expressions of SIGMAR1, BDNF, p-TRKB, the total TRKB, and β-actin across the samples in each group were analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). (E) IF staining of SIGMAR1 (in green), BDNF (in green), and p-TRKB (in red) across the Saline, Meth, SCFA + Saline, and SCFA + Meth samples. Nuclei were labeled with DAPI (in blue). Scale bar, 50 μm. (F) The number of positive cells across each group was shown (n = 6 per group). (G) Spearman correlation analysis of differential microbiota with colonic, behavioral indicators, and IF. The color of the dots denoted the correlation coefficient and the size of the dots denoted the P-value. Red dots represent a positive correlation, while the blue ones represent a negative correlation. Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
Next, we added AA, AP, and their mixture SCFAs into the drinking water of mice respectively to identify which SCFA played a major role in the protective effects (Supporting Information Fig. S9A). In OFT, AA failed to mitigate the Meth-induced decrease in the total travel distance and the time spent in the center zone, and the increase of fecal number (Fig. S9B and S9C). However, Meth-induced deficits of these behavioral indexes (OFT) were effectively attenuated by PA pretreatment and presented no significant difference with the SCFAs + Meth group (Fig. S9B and S9C). In EPMT, the Meth-induced decrease of the time mice spent in open arms was ameliorated by AA, but not PA pretreatment, and both AA and PA significantly alleviated the reduction of the entries mice into open arms (Fig. S9B and S9D). Meanwhile, AA exhibited similar alleviative effects with SCFAs on Meth-induced elevation of immobility time in TST and FST, while the protective effects of PA were not observed (Fig. S9E). Furthermore, the influences of AA, PA, and SCFAs on Meth-induced inhibition of the SIGMAR1/BDNF/TRKB pathway were evaluated. Compared to the Meth group, AA, PA, and SCFAs pretreatment upregulated the expression of proteins SIGMAR1, BDNF, and p-TRKB, while the total TRKB was not altered across the groups. Meanwhile, there was no significant difference in the protein level of SIGMAR1, BDNF, and p-TRKB across the AA + Meth, AP + Meth, and SCFAs + Meth groups (Fig. S9F and S9G). These indicated that the supplementation of AA and PA alone presented the selective improvements of anxiety- and depression-related behavioral indexes, along with the upregulation of SIGMAR1/BDNF/TRKB pathway, while their mixture SCFAs exhibited more comprehensive protection against Meth-related behavioral and neural deficits.
3.5. Sigmar1 knockout disturbed gut microbiome and resulted in similar behavioral phenotypes with Meth exposure by repressing the BDNF/TRKB pathway
Given the regulation of SIGMAR1 on anxiety and depression, the Sigmar1 gene was omitted to investigate its role in Meth-induced mental disorders (Fig. 8A and B).
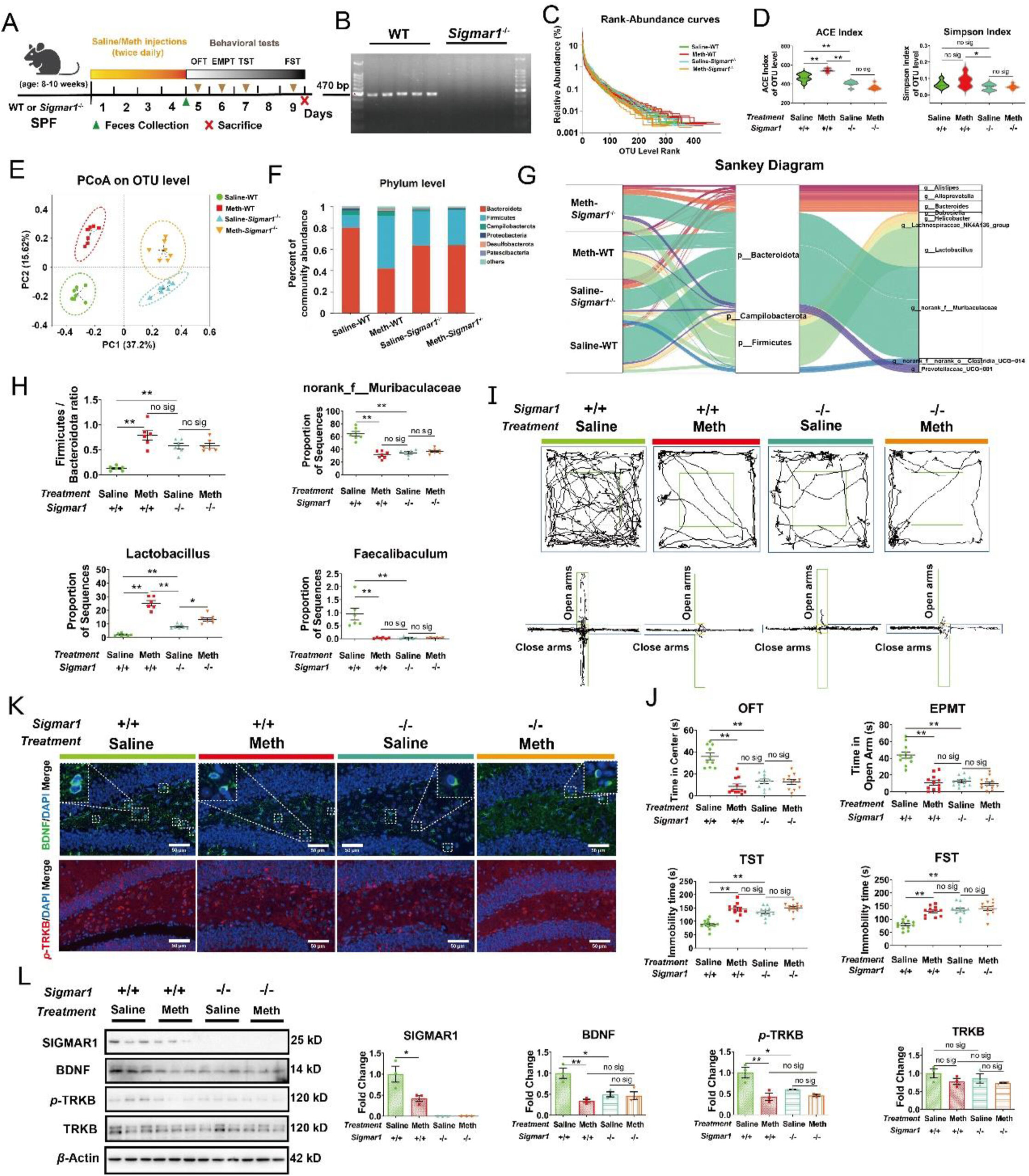
Sigmar1 knockout disturbed the gut microbiome and resulted in similar behavioral phenotypes with Meth exposure.
(A) Schematic diagram of the experimental procedure. (B) Agarose gel electrophoresis of genotype Sigmar1. (C) The Rank-Abundance Curves of all samples. (D) The alteration of alpha diversity across the Saline-WT, Meth-WT, Saline-Sigmar1−/− and Meth-Sigmar1−/− groups, as measured by ACE and Simpson index at the OTU level (n = 7 per group). (E) PCoA plot analysis of the Saline-WT (green dots), Meth-WT (red squares), Saline-Sigmar1−/− (blue positive triangles), and Meth-Sigmar1−/− samples (orange inverted triangles) at the OTU level. (F) Relative abundance of microbiota at the Phylum level. (G) Sankey diagram: relative abundance of differential microbiota at the Phylum level and the corresponding flows of Genus differential microbiota. The color of the ribbon represented the differential microbiota and its width represented the relative abundance of differential microbiota. (H) The ratio of phylum Firmicutes/Bacteroidate across each group (n = 6 per group). The proportion of microbiota sequences for genus norank_f_Muribaculaceae, Lactobacillus, and Faecalibaculum across each group (n = 6–7 per group). Kruskal–Wallis test with a 95% confidence interval was performed to screen the differential microbiota. (I) The representative trajectory diagrams in OFT and EPMT. (J) The time the mice spent in the center zone (OFT, n = 9–12 per group) and the time the mice spent in open arms across each group (EPMT, n = 10 per group). And the total immobility time across each group during the TST and FST (n = 12 per group). (K) IF staining of BDNF (in green) and p-TRKB (in red) across the samples in each group. Nuclei were labeled with DAPI (in blue) (n = 6 per group). Scale bar, 50 μm. (L) Protein expressions of SIGMAR1, BDNF, p-TRKB, the total TRKB, and β-actin across the samples in each group was analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
Interestingly, Sigmar1−/− also presented the regulatory effects on the gut microbiome. Samples among groups showed no significant difference in OTU abundance or evenness (Fig. 8C). In Alpha diversity, Sigmar1−/− mice presented the lower ACE, Chao, and Sobs indexes and Meth failed to increase these indexes, which were different from the observations in WT mice (Fig. 8D; Supporting Information Fig. S10A and S10B). These indicated the lower richness in Sigmar1−/− mice. Meanwhile, in Simpson and Shannon indexes, there was no significant difference observed between Meth and saline administration in both genotypes (Fig. 8D; Fig. S10C). These indicated that Sigmar1−/−reduced the richness but not the diversity of microbiota. In Beta diversity, PCoA analysis displayed that samples from Sigmar1−/− mice were clearly separated from WT mice, and further clustered according to the saline and Meth administration (Fig. 8E). Additionally, the microbial composition of Sigmar1−/− mice was different from the WT mice irrespective of the Meth administration (Fig. 8F; Fig. S10D–S10F). The relative abundance of differential microbiota at the Phylum level and their taxon relationships with differential microbiota at the Genus level in each group were analyzed using the Sankey diagram (Fig. 8G). Specially, we observed that compared with the Sailne-WT group, Sigmar1−/− induced a higher ratio of phylum Firmicutes/Bacteroidate in mice with or without Meth administration and showed no difference with Meth-WT group (Fig. 8H), suggesting the dysbiosis of the intestinal microenvironment in Sigmar1−/− mice. At the Genus level, Saline-Sigmar1−/− mice presented a lower abundance of norank_f_Muribaculaceae, Faecalibaculum, and higher Lactobacillus compared with Saline-WT mice, which were similar to Meth-induced alterations in WT mice (Fig. 8H). Meanwhile, Meth showed no significant influence in norank_f_Muribaculaceae and Faecalibaculum in Sigmar1−/− mice, while Lactobacillus was further increased (Fig. 8H). Thus, we confirmed that SIGMAR1 participated in the regulation of the gut microbiome.
Next, we determined the alterations of the behavioral phenotypes after Sigmar1knockout. The results showed that, in WT mice, Meth resulted in a shorter total travel distance, lesser time spent in the center zone, and greater amount of feces in OFT; the lesser time mice spent in and the entries into the open arms in EPMT; the longer immobility time in TST and FST (Fig. 8I and J; Supporting Information Fig. S11A–S11C). Interestingly, these behavioral alterations were also observed in Sigmar1−/− mice without Meth administration (Fig. 8I and J; Fig. S11A–S11C). In addition, after Sigmar1 knockout, the mice exhibited lower BDNF and p-TRKB expression than Saline-WT mice, regardless of Meth administration (Fig. 8K and L; Fig. S11D). These were similar to the effects of Meth on WT mice. Meanwhile, we did not detect the expression of protein SIGMAR1 in Sigmar1−/− mice, confirming its successful knockout (Fig. 8L). These findings suggested the involvement of SIGMAR1 in Meth-induced depression- and anxiety-like behaviors and that its knockout could cause Meth-like behavioral deficits.
3.6. Sigmar1 knockout eliminated the phenotypic differences in anxiety- and depression-like behaviors between FMT-M and FMT-C mice
To explore if FMT-M-induced depression- and anxiety-like behaviors relied on the modulation of SIGMAR1, we transplanted the microbiota from the saline–or Meth-administrated WT mice into the Sigmar1−/− mice (Fig. 9A). In behavioral tests, compared with the FMT-C-WT group, FMT-C-Sigmar1−/− induced depression- and anxiety-like behaviors, which was demonstrated by the shorter total travel distance, the lesser time spent in the center zone and the greater amount of feces in OFT, the lesser time the mice spent and the fewer entries into the open arms in EPMT, and the longer immobility time in TST and FST (Fig. 9B and C; Supporting Information Fig. S12A–S12C). These suggested that the FMT-M-induced behavioral differences from FMT-C in WT mice were leveled out by Sigmar1 knockout. Additionally, FMT-C-Sigmar1−/− displayed a lower level of BDNF and p-TRKB than the FMT-C-WT mice and showed no significant difference with the FMT-M-Sigmar1−/− mice (Fig. 9D and E; Fig. S12D and S12E). These results suggested that the regulation of the gut microbiota on depression and anxiety relied on the modulation of the SIGMAR1/BDNF/TRKB pathway.

I) Sigmar1 knockout eliminated the phenotypic differences in behaviors between FMT-M and FMT-C mice.
(A) Schematic diagram of the experimental procedure. (B) The representative trajectory diagrams in OFT and EPMT. (C) The time the mice spent in the center zone (OFT, n = 10–11 per group) and the time the mice spent in open arms across each group (EPMT, n = 10–12 per group). The total immobility time across each group during TST and FST. n = 12 per group. (D) IF staining of BDNF (in green) and p-TRKB (in red) across the samples in each group. Nuclei were labeled with DAPI (in blue) (n = 6 per group). Scale bar, 50 μm. (E) Protein expressions of BDNF, p-TRKB, the total TRKB, and β-actin across the samples in each group were analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). II) Sigmar1 knockout eliminated the protective effects of SCFAs on Meth-induced depression- and anxiety-like behaviors. (F) Schematic diagram of the experimental procedure. (G) The representative trajectory diagrams in OFT and EPMT. (H) The time the mice spent in the center zone (OFT, n = 10–11 per group) and the time mice spent in the open arms (EPMT, n = 10–12 per group) across each group. The total immobility time across each group during TST and FST (n = 12 per group). (I) IF staining of BDNF (in green) and p-TRKB (in red) across the samples in each group. Nuclei were labeled with DAPI (in blue) (n = 6 per group). Scale bar, 50 μm. (J) Protein expressions of BDNF, p-TRKB, the total TRKB, and β-actin across the samples in each group was analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
3.7. Sigmar1 knockout abolished the protective effects of SCFAs on Meth-induced anxiety- and depression-like behaviors by suppressing the BDNF/TRKB pathway
Next, we investigated the role of SIGMAR1 in the protection of SCFAs on Meth-induced mental disorders (Fig. 9F). Behavioral tests showed that there was no significant difference in depression- and anxiety-related indicators between the SCFA-Meth-Sigmar1−/− and Meth-WT groups, except for the time spent in the center zone (OFT) (Fig. 9G and H; Supporting Information Fig. S13A–S13C), suggesting that SCFAs failed to mitigate Meth-induced depression- and anxiety-like behaviors after Sigmar1 knockout. Moreover, SCFAs failed to attenuate the Meth-induced decrease of BDNF and p-TRKB in Sigmar1−/− mice (Fig. 9I and J; Fig. S13D and S13E), which were different from the observations in WT mice. Thus, our results indicated that the protective effects of SCFAs on Meth-induced depression- and anxiety-like behaviors were dependent on the modulation of the SIGMAR1/BDNF/TRKB pathway.
3.8. Fluvoxamine, an agonist of SIGMAR1, alleviated Meth-induced anxiety- and depression-like behaviors by activating the SIGMAR1/BDNF/TRKB pathway
To verify the protection of SIGMAR1 on Meth-induced mental disorders, Flu was utilized to activate the SIGMAR1 (Fig. 10A). In behavioral tests, we observed that the Meth-induced depression- and anxiety-like behaviors were also attenuated by Flu treatment, as demonstrated by the longer total travel distance, the more time spent in the center zone, and the lower amount of feces in OFT, the longer time the mice spent and the more entries into the open arms in EPMT, and the shorter immobility time in TST and FST after Flu treatment compared with the Meth group (Fig. 10B and C; Supporting Information Fig. S14A–S14C). Furthermore, compared to the Saline group, we observed that we observed that Flu overexpressed the level of SIGMAR1 in both saline- and Meth-administrated mice and attenuated the Meth-induced downregulation of BDNF and p-TRKB (Fig. 10D and E; Fig. S14D). These observations further confirmed the protective effects of SIGMAR1 on Meth-induced depression- and anxiety-like behaviors.
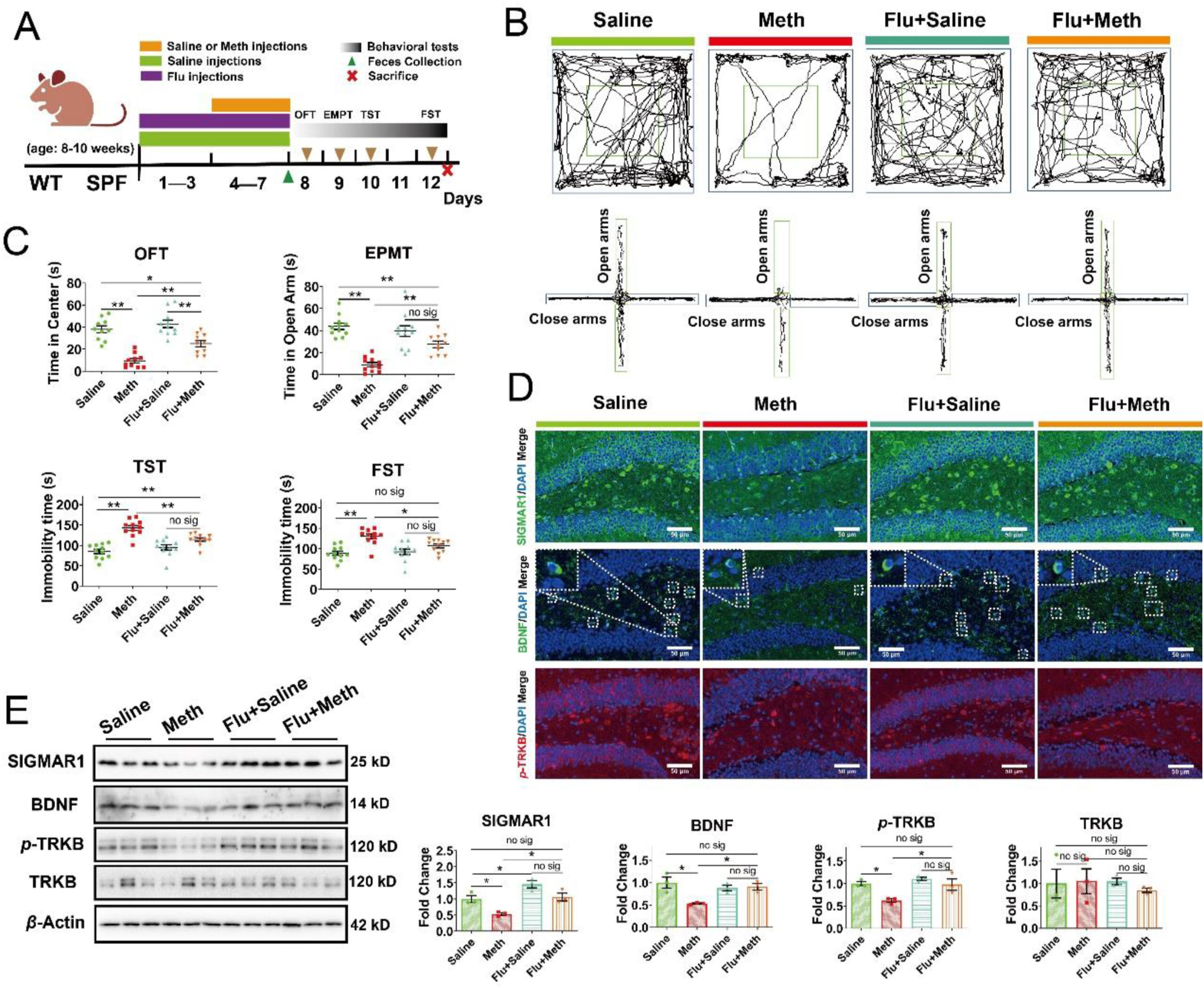
Fluvoxamine ameliorated Meth-induced anxiety- and depression-like behaviors by stimulating the SIGMAR1/BDNF/TRKB pathway.
(A) Schematic diagram of the experimental procedure. (B) The representative trajectory diagrams in OFT and EPMT. (C) The time the mice spent in the center zone (OFT, n = 10–12 per group) and the time the mice spent in open arms (EPMT, n = 10–11 per group) across each group. The total immobility time across each group during TST and FST (n = 11 per group). (D) IF staining of SIGMAR1 (in green), BDNF (in green), and p-TRKB (in red) across the Saline, Meth, Flu + Saline, and Flu + Meth samples. Nuclei were labeled with DAPI (in blue) (n = 6 per group). Scale bar, 50 μm. (E) Protein expressions of SIGMAR1, BDNF, p-TRKB, the total TRKB, and β-actin across the samples in each group was analyzed by Western blot. β-Actin served as a loading control (n = 3 per group). Data are expressed as mean ± SEM, ∗P < 0.05, ∗∗P < 0.01, no sig: no significant difference (P > 0.05). All experiments were performed in triplicate at a minimum.
Next, we profiled the alteration of gut microbiome after Flu treatment. Rank-Abundance curves presented similar OTU abundance or evenness among all samples (Supporting Information Fig. S15A). In Alpha diversity, Meth induced a higher ACE index than the Saline group, while this alteration was mitigated by Flu treatment (Fig. S15B). Meanwhile, there was no significant difference between the Saline and Meth groups, while Flu + Meth resulted in the elevation of the Simpson index relative to the Saline group (Fig. S15B). These suggested that Flu could ameliorate the Meth-induced richness of microbiota and induce a higher microbial diversity. In Beta diversity, we observed that the principal composition of fecal samples in the Saline group was separated from the Meth group, and Flu altered the sample composition of both groups (Fig. S15C). Moreover, the abundance of gut microbiota from Phylum to Genus level was altered by Flu treatment (Fig. S15D–S15F). The differential microbiota from Phylum to Genus level were further summarized using the LEfSe cladogram (Fig. S15G). In addition, the concentration of SCFAs in Flu-treated fecal samples was also determined (Fig. S15H). Interestingly, mice treated with Flu showed a much higher level of acetic acid, hexanoic acid, and isohexanoic acid than those without Flu treatment, regardless of Meth administration (Fig. S15H). Meanwhile, Flu attenuated the Meth-induced decrease of propanoic acid, while the lowered isobutyric acid and isovaleric acid level was not effectively alleviated by Flu treatment. And there was no significant difference in the concentration of Butanoic acid and Valeric acid across the groups (Fig. S15H). These results indicated that Flu treatment could reshape Meth-induced disturbance of gut microbiota and partly ameliorate the decrease of fecal SCFAs. Moreover, we explored the influences of Flu on the level of serum SCFAs. The results showed that Meth led to a decrease in serum acetic acid, propanoic acid, and hexanoic acid compared to the Saline group, while Flu treatment did not mitigate these changes (Fig. S15I). Additionally, there were no significant differences in the levels of serum butanoic acid, valeric acid, isobutyric acid, and isovaleric acid across the groups (Fig. S15I). As for the level of isohexanoic acid, the Flu + Meth group exhibited a lower concentration than the Saline group (Fig. S15I). These observations suggested that Flu exerted only a weak regulatory influence on Meth-induced alterations in serum SCFAs composition.
Finally, to investigate the role of microbiota in Flu-mediated protective effects, mice were orally pretreated with fecal microbiota from Flu-treated mice before Meth challenges (Supporting Information Fig. S16A). Behavioral results showed that in OFT, there was no significant difference in the total travel distance, time mice spent in the center zone, and the fecal number between the FMT-Flu + Meth and PBS + Meth groups (Fig. S16B and S16C). In EPMT, gut microbiota from Flu-treated mice failed to ameliorate Meth-induced decrease in the time mice spent in and the entries into open arms (Fig. S16B and S16D). Meanwhile, in TST and FST, FMT-Flu + Meth also presented no significant difference in immobility time with the PBS + Meth group (Fig. S16E). In addition, gut microbiota from Flu-treated mice also failed to attenuate Meth-induced repression of the SIGMAR1/BDNF/TRKB pathway. FMT-Flu + Meth resulted in a lower level of proteins SIGMAR1, BDNF, and p-TRKB in comparison with the PBS + Saline group and presented no significant difference with the PBS + Meth group (Fig. S16F and S16G). These findings suggested that Flu-induced alteration of gut microbiota did not contribute to the protective effects of Flu on Meth-induced depression- and anxiety-like behaviors, and the upregulation of the SIGMAR1/BDNF/TRKB pathway.
4. Discussion
Gut microbiota is a crucial contributor to maintaining the host health57. Its-derived SCFAs are recognized as the important mediums of gut microbiota to regulate the gut immune and to influence the mental health of the host20, 21, 22, 23. In this study, the inflammation-related TLR4/NF-κB pathway was significantly enriched after Meth administration based on RNA-seq. Meanwhile, the anti-inflammatory bacterium Faecalibaculum58 was also lowered by Meth, implying that microbial dysbiosis could participate in Meth-induced colonic inflammation. Correspondingly, we observed that gut microbiota from Meth-administrated mice also aroused the TLR4-related colonic inflammation and caused the dysfunction of the intestinal barrier in recipients. TLR4 is a member of the TLR family inducing pro-inflammation response and can be activated by LPS (the component of Gram-negative bacteria)59, while the pro-inflammatory effect of microbial dysbiosis is the important avenue to impair the intestinal barrier function60. The higher serum LPS level was also observed after FMT-M administration. Moreover, the decrease in SCFAs could be a trigger factor for these consequences. In fact, SCFAs can serve as ligands for G-protein coupled receptors, which further activate anti-inflammatory signaling cascades61,62. Additionally, SCFAs have emerged with the capacity of improving the intestinal mucosal barrier62. Indeed, SCFAs supplementation effectively ameliorated Meth-induced colonic inflammation by suppressing the TLR4/NF-κB pathway and strengthening the intestinal barrier. These findings confirmed that gut microbiota participated in Meth-induced impairment of intestinal homeostasis, which could be mitigated by SCFAs. Furthermore, accumulating evidence suggests that there exhibited complex interactions between colonic inflammation and mental disorders, including depression and anxiety63. Patients with inflammatory bowel diseaseexperienced a higher risk of depression and anxiety. And in experimental models of colitis, rodents also presented strong depression- and anxiety-like behaviors64, 65, 66. Accompanied by the colonic inflammation, Meth also caused depression- and anxiety-like behaviors in mice, implying that colonic inflammation could contribute to Meth-related mental disorders. Additionally, behavioral phenotypes of depression and anxiety in inflammatory bowel disease patients could be transferred to the recipient mice by fecal microbiota transplantation67. Here, the FMT experiment reproduced Meth-induced depression- and anxiety-like behaviors in recipients, suggesting that gut microbiota could serve as the linker between Meth-related colonic inflammation and mental disorders.
Notably, Meth induced opposed alterations of mainly SCFAs producers: the downregulated Muribaculaceae and upregulated Lactobacillaceae68. We speculated that it could involve in the microbial mechanism of rescue. Muribaculaceae was the dominant bacteria and its downregulation reduced the SCFAs level, while the increased Lactobacillaceae failed to reverse it. Correspondingly, SCFAs supplementation normalized the abundance of Lactobacillaceae and repressed the recovery of Muribaculaceae, which prevented the overproduction of SCFAs. In addition, probiotic Lactobacillaceae can promote the proliferation of goblet cells to improve the function of the intestinal barrier69,70. Here, accompanied by the upregulated Lactobacillaceae, we also observed the excessive proliferation of goblet cells after Meth administration, although the barrier function was not ameliorated.
In addition, Meth-induced anxiety- and depression-like behaviors were transferred to the recipients through fecal microbiota transplant, indicating the involvement of microbiota in Meth-induced anxiety and depression. An increasing body of evidence presented the regulatory effects of gut microbiota on mental disorders9, 10, 11. Gut microbiota was observed disordered in depression patients, and its transplant induced depression-like behavior of germ-free mice10,12. Even gene-mediated anti-anxiety and -depression phenotypes could be transferred through fecal microbiota transplant71. Furthermore, accompanied by these behavioral phenotypes, we observed the repression of the SIGMAR1/BDNF/TRKB pathway both after Meth and FMT-M administration, implying its potential regulation of behavioral deficits. In fact, the pharmacological activation of SIGMAR1 has been recognized as an effective therapeutic strategy for anxiety and depression in clinic33. Meanwhile, its downstream BDNF and TRKB are also the key molecules for anti-anxiety and -depression35, which could be modulated by gut microbiota72,73. Further, through Sigmar1 knockout and activation, we verified that Meth-induced anxiety and depression relied on the inhibition of SIGMAR1/BDNF/TRKB pathway. In parallel, Sigmar1 knockout eliminated microbiota-induced differences in behavioral phenotypes, confirming that SIGMAR1 mediated microbiota-induced anxiety- and depression-like behaviors. Additionally, we observed that Sigmar1 knockout also disturbed the gut microbiome. In our previous study, gut microbiota from Sigmar1knockout mice also partly reproduced genotype-related depression- and anxiety-like behaviors74. These suggested that there existed a dynamic balance between gut microbiota and SIGMAR1, and gut microbiota could contribute to the modulation of SIGMAR1 on behaviors.
In this study, we observed a decrease in microbiota-derived SCFAs, both in feces and serum, due to Meth administration. Supplementation with SCFAs alleviated Meth-induced depression- and anxiety-like behaviors, potentially through regulation of the SIGMAR1/BDNF/TRKB pathway. Through oral supplementation, SCFAs could regulate the homeostasis of colonic Treg cells75, suggesting that oral SCFAs supplementation could transit through the small intestine and act on the colonic tissue. Meanwhile, both oral SCFAs supplementation and microbial colonization could improve microbial knockout-induced deficiency of microglia in the brain76, indicating that gut microbiota and its derivation SCFAs played a crucial role in modulating neural function. In fact, the protective effects of SCFAs were observed in diet-related depression77 and even in chronic cerebral hypoperfusion-induced depression78. SCFAs also presented the alleviative effects on innate anxiety79 and the anti-anxiety effects of prebiotics strongly correlated with gut SCFAs level25. In addition, SCFAs failed to ameliorate Meth-induced anxiety and depression in Sigmar1−/− mice, indicating that the protective effects of SCFAs against Meth-induced mental disorders were dependent on the modulation of SIGMAR1.
By activating the SIGMAR1, Flu upregulated the BDNF/TRKB pathway and ameliorated Meth-induced depression- and anxiety-like behaviors. This further confirmed that Meth-induced mental disorders relied on its repression of SIGMAR1. Interestingly, we also observed that Flu treatment reshaped Meth-induced microbial disturbance and partly attenuated Meth-induced decrease of SCFAs, implying that gut microbiota could participate in the protective effects of Flu. However, gut microbiota from Flu-treated mice failed to alleviate Meth-induced inhibition of the SIGMAR1/BDNF/TRKB pathway and showed no effective protection against depression- and anxiety-like behaviors. Flu exhibited a limited regulatory effect on Meth-induced alterations in serum SCFAs. These findings suggest that the gut microbiota-mediated short-term elevation of fecal SCFAs was insufficient to counteract the Meth-induced decrease in serum SCFAs or to alleviate associated mental disorders (as the SCFAs supplementation experiment lasted three weeks). Furthermore, Flu exerted its antidepressant and anxiolytic effects primarily by directly targeting SIGMAR1.
However, there are still a few shortcomings of this study to be noted. Here, the detailed mechanisms of how SCFAs modulate Meth-induced anxiety and depression disorders are still unclear, especially the potential regulation of free fatty acid receptor on SIGMAR1. Another shortcoming is that our study only focused on the protective effects of acetic acid and propanoic acid, while the role of other SCFAs needs to be further clarified.
5. Conclusions
Our findings indicate that gut microbiota participated in Meth-induced colonic inflammation and contributed to Meth-related anxiety and depression disorders; microbiota-derived SCFAs could optimize gut homeostasis and protect against Meth-induced anxiety and depression in SIGMAR1 dependent manner. Overall, this study revealed the microbial mechanisms of Meth-induced anxiety and depression disorders and provided a potential candidate in therapeutic strategy.