
Activation of σ-1 receptor mitigates estrogen withdrawal-induced anxiety/depressive-like behavior in mice via restoration of GABA/glutamate signaling and neuroplasticity in the hippocampus

By Peng Ren, Jing-Ya Wang, Hong-Lei Chen, Yue Wang, Lin-Yu Cui, Jing-Yao Duan, Wen-Zhi Guo, Yong-Qi Zhao, and Yun-Feng Li
Excerpt from the article published in Journal of Pharmacological Sciences, Volume 154, Issue 4, April 2024, Pages 236-245, ISSN 1347-8613, DOI: https://doi.org/10.1016/j.jphs.2024.02.003.
Abstract
Postpartum depression (PPD) is a significant contributor to maternal morbidity and mortality. The Sigma-1 (σ-1) receptor has received increasing attention in recent years because of its ability to link different signaling systems and exert its function in the brain through chaperone actions, especially in neuropsychiatric disorders. YL-0919, a novel σ-1 receptor agonist developed by our institute, has shown antidepressive and anxiolytic effects in a variety of animal models, but effects on PPD have not been revealed. In the present study, excitatory/inhibitory signaling in the hippocampus was reflected by GABA and glutamate and their associated excitatory-inhibitory receptor proteins, the HPA axis hormones in the hippocampus were assessed by ELISA. Finally, immunofluorescence for markers of newborn neuron were undertaken in the dentate gyri, along with dendritic spine staining and dendritic arborization tracing. YL-0919 rapidly improves anxiety and depressive-like behavior in PPD-like mice within one week, along with normalizing the excitation/inhibition signaling as well as the HPA axis activity. YL-0919 rescued the decrease in hippocampal dendritic complexity and spine density induced by estrogen withdrawal. The study results suggest that YL-0919 elicits a therapeutic effect on PPD-like mice; therefore, the σ-1 receptor may be a novel promising target for PPD treatment in the future.
1. Introduction
Postpartum depression (PPD) is the most common psychiatric disorder following childbirth, and is characterized by maternal mood disturbance, reduced interest or pleasure in matters, and excessive concern or indifference to infants,1 all of which have a negative impact on offspring development.2 The worldwide prevalence of PPD is approximately 15%; the prevalence of PPD varies from region to region due to differences in economic levels and screening awareness, and can be underestimated.3 Like many psychiatric disorders, PPD likely has biological and psychosocial components. Indeed, PPD has unique features involving dysregulation of the hypothalamic–pituitary–adrenal (HPA) stress axis, peptide and steroid hormones, and GABA circuitry during pregnancy. Increasing evidence has demonstrated dramatic fluctuations in steroid hormones (i.e., progesterone, allopregnanolone) during pregnancy and the postpartum period,4 and these alterations result in an abnormal GABAergic signaling,5 which may exacerbate the symptoms in vulnerable mothers.
Under- or un-treated PPD can lead to multiple short- and long-term adverse outcomes for mothers, infants, and families, and sudden or severe adverse outcomes, such as suicide and infanticide, indicate the need for rapid resolution of PPD symptoms.6 Despite the unique physiology associated with pregnancy and childbirth, many similarities interact the mechanisms of disease for PPD and major depressive disorder (MDD). The current first-line antidepressant medications for PPD treatment are the selective serotonin reuptake inhibitors (SSRIs).7 While often effective, newborns of mothers treated with SSRIs may be at risk of adverse perinatal outcomes including withdrawal symptoms and cardiac malformation.8 In 2019, Brexanolone (SAGE-547), the first drug approved by the Food and Drug Administration for the treatment of PPD, has been shown to rapidly reduce PPD symptoms.9,10 However, this new drug, which brings hope to many new mothers suffering from depression, is not only expensive, but also has serious side effects including loss of consciousness and excessive sedation,10 which have limited its widespread use in the clinic. Therefore, understanding the pathogenesis of PPD action is important for further development of cost-effective ideal medications with minimal side effects.
The σ-1 receptor, a type of chaperone protein, has attracted increasing attention recently due to its multiple cellular functions.11,12 It is mainly located in the cholesterol-rich region of the endoplasmic reticulum (ER) and mitochondria-associated membranes and is highly expressed in brain regions involved in emotion and neuropsychiatric disorders,13 especially in astrocytes.14 One functional change in reactive astrocytes observed in animal models of mental disease involves the biosynthesis of glutamate (Glu) and GABA, followed by the extracellular release of these neurotransmitters, which can mediate neuronal excitation or inhibition.15,16 It is already known that many currently available antidepressants bind to σ-1 receptors; therefore, their pharmacological effects may depend partially on these receptors.17 In addition, some endogenous neurosteroids, such as progesterone, dehydroepiandrosterone, etc., are reported to be endogenous ligands or positive allosteric regulators for σ-1 receptors,18and are also involved in regulating anxiety and depression in PPD.2,19,20 Therefore, this suggests that the σ-1 receptor may also be involved in PPD molecular mechanisms, although this is rarely reported in the literature.
YL-0919 [(1-(1-benzyl-4-hydroxypiperidin-4-ylmethyl)-2(1H)-pyridinone hydrochloride], is a novel compound developed in our institute, which can rapidly and effectively alleviate anxiety- and depressive-like behaviors in a variety of animal models,21,22,23 and is currently being identified as a selective σ-1 receptor agonist.12Evidence from clinical and biological studies suggests that the hippocampus is highly sensitive to stress and its dysfunction plays an important role in the development of depression.24,25 In short, this study aimed to evaluate the pharmacological effects of YL-0919 on PPD-like mice and further clarification of σ-1 receptor as one of the crucial pharmacological targets of YL-0919 fast-acting antidepressants. An additional aim was to study the possible mechanism of action involving the σ-1 receptor in the PPD-like mouse hippocampus, providing a fundament for the development of novel antidepressants targeting on σ-1 receptor.
..
3. Results
3.1. YL-0919-mediated relief of anxiety- and depressive-like behaviors in PPD-like mice
After 6 days of drug administration, mice sequentially experienced the behavioral tests. In the OFT (Fig. 1 B), there is no significant difference in the total moved distance across groups (Fig. 1C left). Notably, obvious anxiety-like behaviors in mice were induced by estrogen withdrawal, including reduced entries in the central zone of the OFT (F (3, 56) = 3.901, p < 0.05, Fig. 1C right), as well as a decreased during time in the central area of the EPM (F (3, 55) = 2.452, p < 0.05, Fig. 1D left) and the percentage of entries in the open arms (F (3, 56) = 3.524, p < 0.05, Fig. 1D right). However, the anxiety-like behaviors in PPD-like mice were alleviated by PRE-084 or YL-0919 treatment (OFT: PRE-084, p < 0.05, YL-0919, p < 0.05; EPM: during time in the central area, PRE-084, p < 0.05; percentage of entries in the open arms, YL-0919, p < 0.05, Fig. 1D). Furthermore, the selective σ-1 receptor antagonist, NE-100 (0.5 or 1.0 mg/kg) administration totally reversed the anxiolytic effect of YL-0919 observed in the OFT (F (2, 42) = 3.720, NE-100, 1.0 mg/kg, p < 0.05, Fig. 1C right) and EPM (percentage of entries in the open arms: F (2, 42) = 5.860, NE-100, 0.5 mg/kg, p < 0.01, Fig. 1D right). These data suggest that the σ-1 receptor plays a core role in regulating the anxiolytic effect in PPD-like mice treated with YL-0919. Next, the depression-like behaviors in PPD-like mice were assessed with the SPT, TST, and FST tests. The sucrose preference of the OVX-vehicle group mice was significantly decreased compared to the non-OVX mice (F (3, 56) = 6.563, p < 0.01, Fig. 1E). Similarly, the results of the TST (F (3, 54) = 2.956, p < 0.01, Fig. 1F) and FST (F (3, 56) = 3.923, p < 0.01, Fig. 1G) showed that immobility time were markedly increased in the OVX-vehicle group mice compared to the non-OVX-vehicle group. The congruent results provide strong evidence that the PPD-like mice exhibit significant depressive-like behaviors. After 1 week of treatment, YL-0919 restored the sucrose preference (p < 0.01, Fig. 1E) and significantly decreased the immobility time in the FST of the PPD-like mice (p < 0.05, Fig. 1G). Interestingly, the effects of YL-0919 were blurred by NE-100 (SPT: F (2, 42) = 4.739, p < 0.01; FST: F (2, 42) = 4.031, p < 0.05), which indicates that YL-0919 acts through the activation of σ-1 receptors.
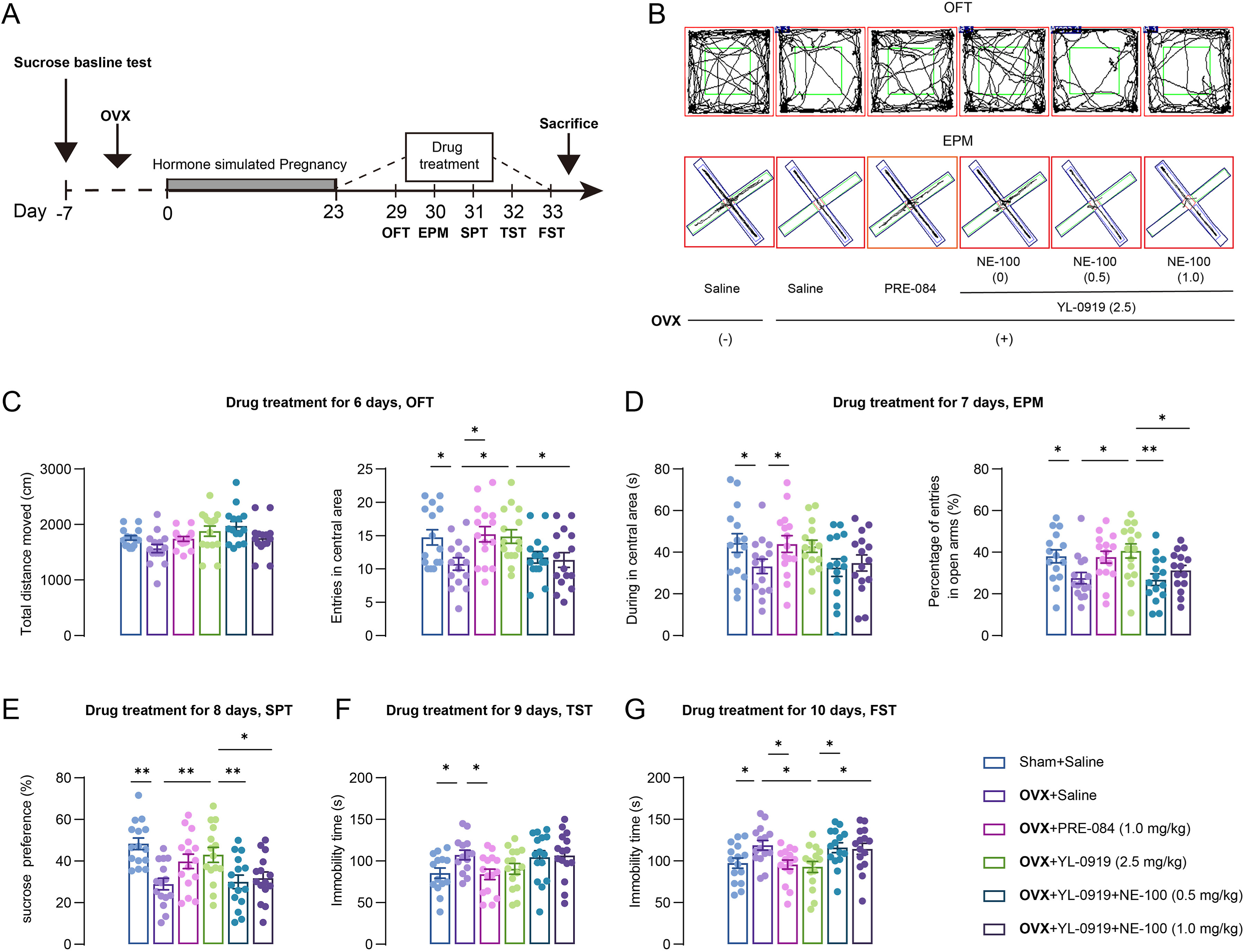
YL-0919 mitigated the anxiety- and depression-like behaviors in PPD-like mice.
(A) The time flowchart of hormone-simulated pregnancy (HSP), drug treatment, and behavioral experiments. (B) Representative trace recordings of mice in the open-field test (OFT) (top) and the elevated plus maze (EPM) (bottom). (C) The total travel distance (left) and entries in the central area (right) over a 5-min period in the OFT were detected. (D) The duration of time in the central area (left) and the percentage of entries into the open arms (right) in the EPM. (E) The effects of drug treatment on the sucrose preference of mice. The immobility time of mice in the tail suspension test (TST) (F) and the forced swim test (FST) (G). Data are presented as mean ± SEM, (n = 14–15/group), *p < 0.05, **p < 0.01.
3.2. Effects of YL-0919 on Glu/GABA levels in PPD-like mouse hippocampal astrocytes
Glu and GABA are the main excitatory and inhibitory neurotransmitters respectively, in the central nervous system. Here we detected the Glu and GABA levels in PPD-like mouse hippocampal astrocytes. Expression of Glu and GABA in astrocytes was determined by the regions occupied by Glu and GABA in GFAP-positive cells. The results reveal that the GABA levels in PPD-like mice were decreased in comparison with non-OVX mice (CA1: F (3, 19) = 3.919, p < 0.01; CA3: F (3, 19) = 4.362, p < 0.01, Fig. 2A and B), whereas the Glu levels were upregulated in CA1 region (F (3, 20) = 4.063, p < 0.01, Fig. 2C and D). Following YL-0919 treatment, their levels were reversed (GABA: CA3, p < 0.05; Glu: CA1, p < 0.05), similar with treatment of the positive control drug PRE-084 (GABA, CA1: p < 0.05; Glu, CA1, p < 0.05). However, the effect of YL-0919 was abolished by NE-100 treatment (GABA: F (2, 12) = 2.788, p < 0.05; Glu: F (2, 15) = 3.057, p < 0.05, Fig. 2).
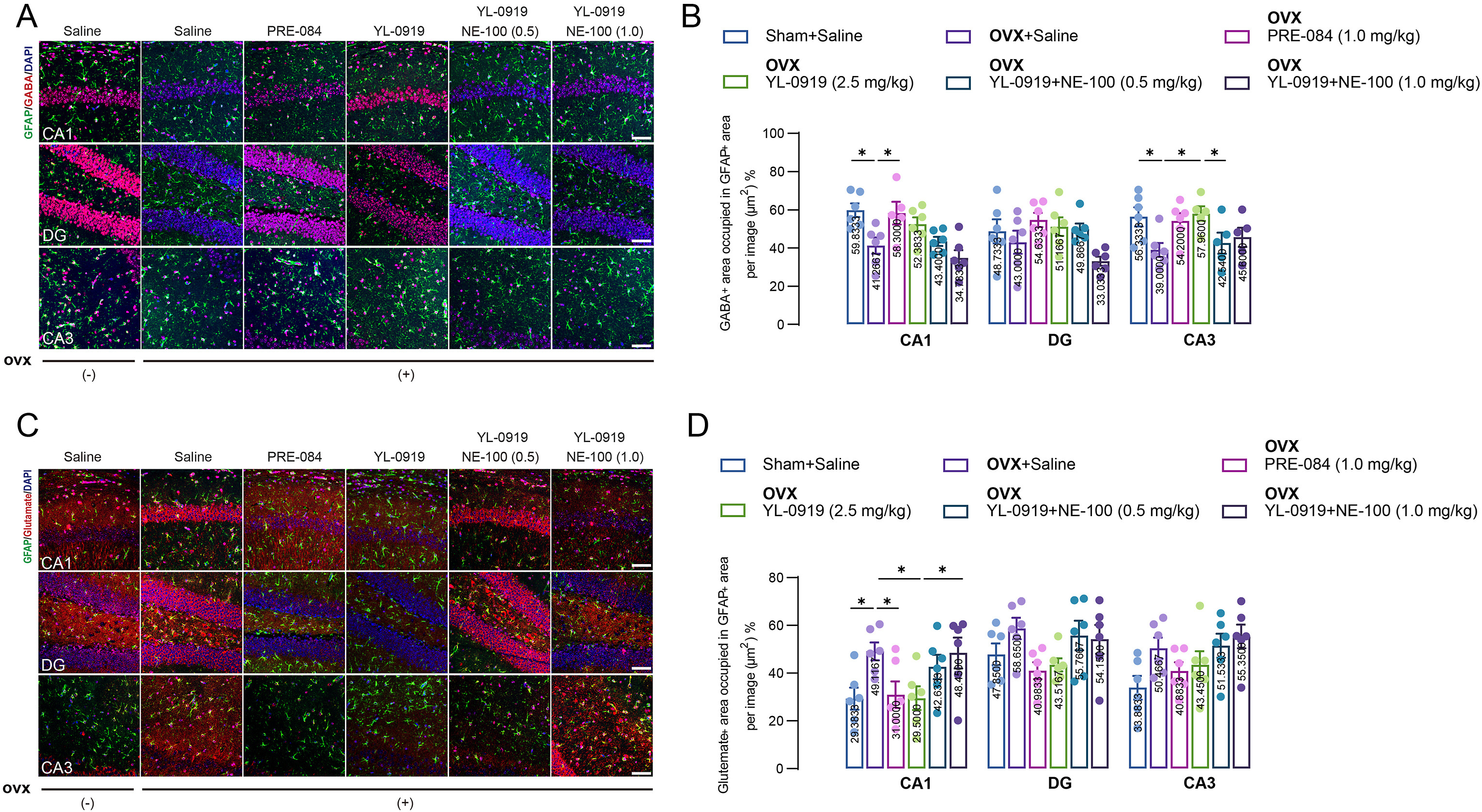
YL-0919 regulates Glu/GABA levels in PPD-like mouse hippocampal astrocytes.
(A) Representative micrographs of GFAP (green) and GABA-positive (red) cells in hippocampal CA1, DG, and CA3 regions. (B) The intensity of GABA+ GFAP+ was quantified in each group. (C) Representative micrographs of GFAP (green) and Glu-positive (red) cells in hippocampal CA1, DG, and CA3. (D) The intensity of GFAP+ Glu+ was quantified in each group. Scale bar = 50 μm. Data are presented as mean ± SEM (n = 5–6 from three mice each group), *p < 0.05, **p < 0.01. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
3.3. YL-0919 activates σ-1 receptor and regulates the excitation/inhibition related protein expression in the hippocampus of PPD-like mice
It has been suggested that perturbations in the excitation/inhibition balance within the brain can result in an array of neuropathological and psychiatric aberrations, encompassing conditions such as schizophrenia, attention deficit hyperactivity disorder, anxiety, and depression.34 We detected the alterations in the corresponding receptors for the above mentioned two types of transmitters. YL-0919 treatment activated the σ-1 receptor and upregulate its expression (F (3, 20) = 5.673, p < 0.05, Fig. 3A–D), correspondingly the level of phosphorylation GluA1 at the Ser845 site (p-GluA1-Ser845) was decreased in the hippocampus of drug treated PPD-like mice (F (3, 20) = 4.544, PRE-084: p < 0.05, YL-0919: p < 0.01, Fig. 3B–E). In addition, the effect of YL-0919 was abolished by NE-100 treatment (σ-1 receptor: F (2, 15) = 3.217, p < 0.05; p-GluA1: F (2, 15) = 2.438, p < 0.05). Further results revealed that the GABAergic receptors, including the representative ionic receptor GABA(A)R1 and metabolic receptor GABA(B)R1/2, were decreased in PPD-like mice (GABA(A)R1: F (3, 20) = 6.817, p < 0.01; GABA(B)R1: F (3, 20) = 3.433, p < 0.05; GABA(B)R2: F (3, 20) = 3.173, p < 0.01, Fig. 3F–H), whereas PRE-084 or YL-0919 treatment upregulated their expression levels. NE-100 treatment reversed the abovementioned effects of YL-0919 (GABA(B)R1: P = 0.0470, p < 0.05; GABA(A)R1: F (2, 15) = 5.894, p < 0.01, Fig. 3C, F–H).
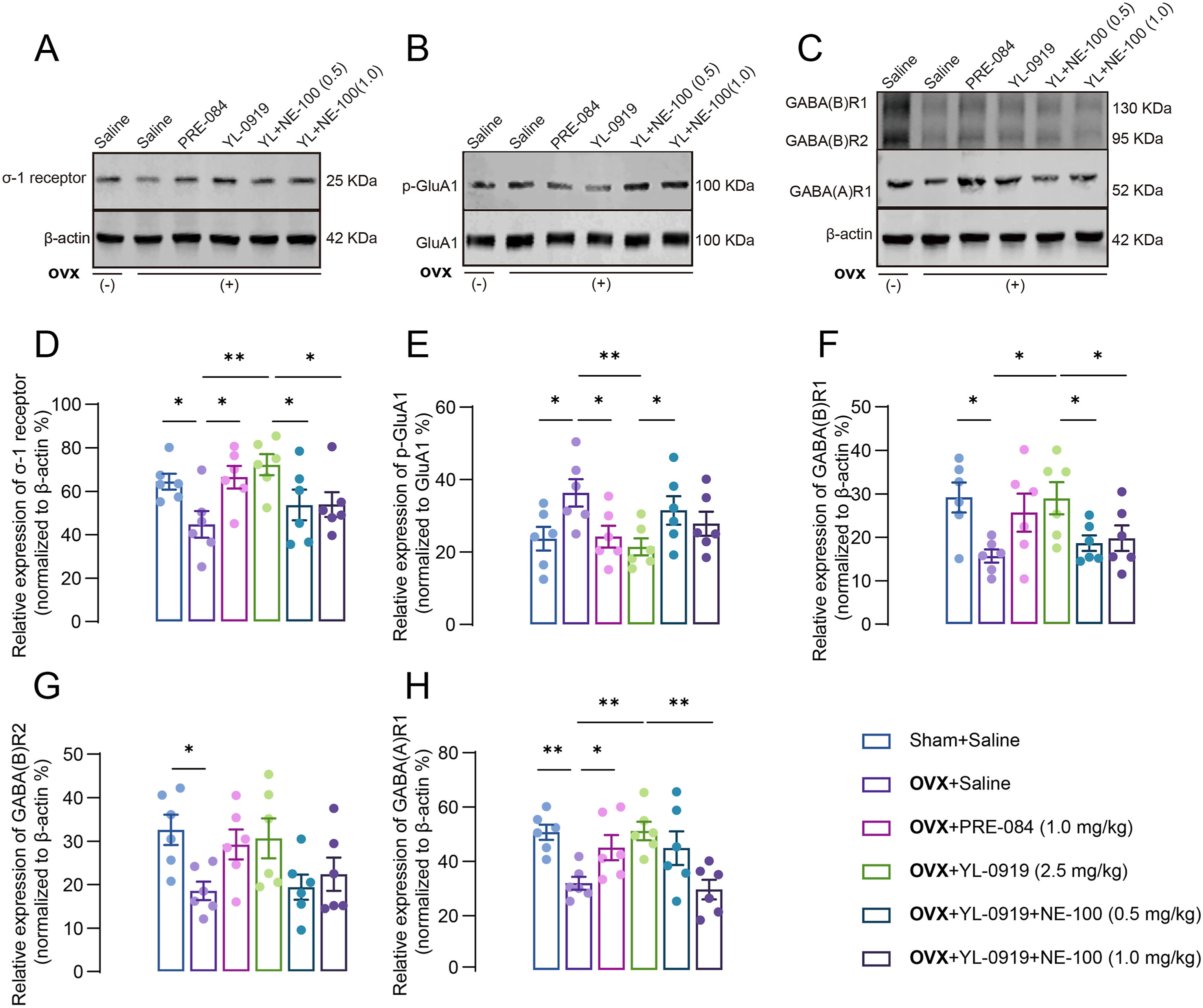
The effect of YL-0919 on the excitation/inhibition signal in the hippocampus of PPD-like mice.
The representative Western blot for the σ-1 receptor (A), p-Glua1/Glua1 (B) and GABA(A)R1 and GABA(B)R1/2 (C) shown in the hippocampus. Quantification of the gray value of each group representative blot image, σ-1 receptor (D), p-Glua1 (E), GABA(B)R1/2 (F–G), and GABA(A)R1 respectively. Data are presented as mean ± SEM, (n = 6 from three mice each group), *p < 0.05, **p < 0.01.
3.4. Effects of YL-0919 on HPA axis hormones in PPD-like mice
HPA axis hormones are involved in the occurrence and development of PPD2; therefore, we measured their levels in the serum of PPD-like mice. The levels of ACTH and CORT were obviously increased (ACTH: F (3, 20) = 6.131, p < 0.05; CORT: F (3, 20) = 5.341, p < 0.01, Fig. 4A and B) in the serum of PPD-like mice as compared to those of non-OVX mice. PRE-084 or YL-0919 treatment significantly reverted the ACTH and CORT concentrations to basal levels; however, NE-100 treatment abolished the effects of YL-0919 (ACTH: F (2, 15) = 5.473, p < 0.01; CORT: F (2, 15) = 4.957, p < 0.01, Fig. 4A and B).
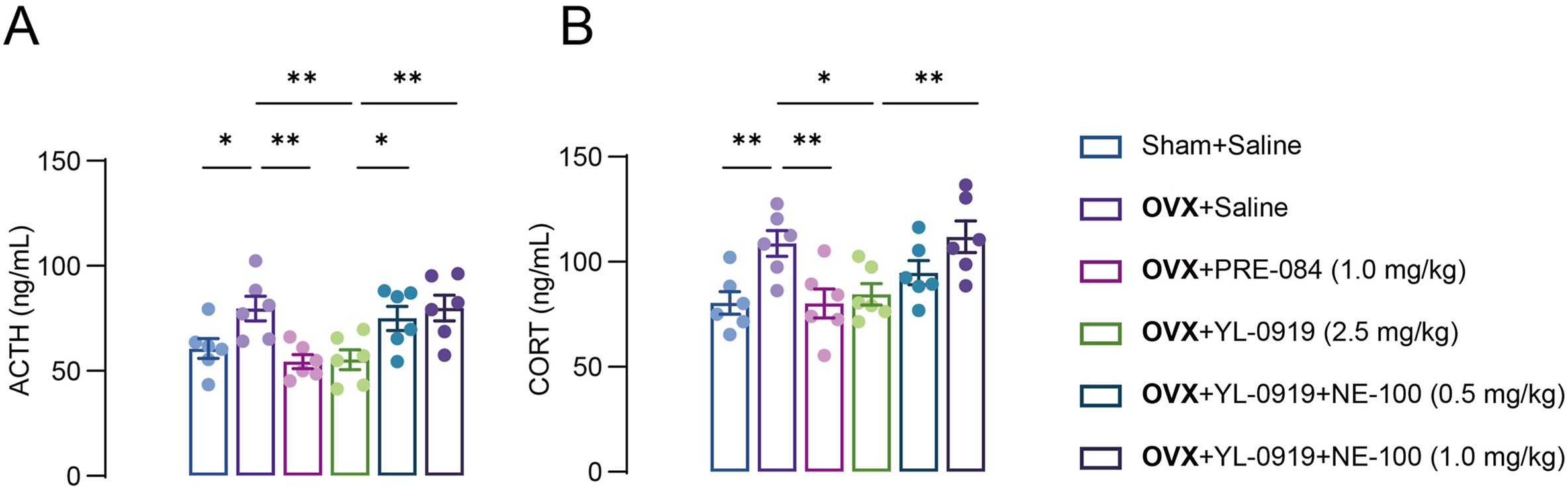
YL-0919 regulates the HPA axis hormone levels in the PPD-like mice.
The adrenocorticotropic hormone (ACTH) (A) and corticosterone (CORT) (B) levels were significantly increased in the sera of PPD-like mice. YL-0919 or PRE-084 restored the ACTH and CORT concentration to basal levels, which were blocked by NE-100 treatment. Data are presented as mean ± SEM, (n = 6/group), *p < 0.05, **p < 0.01.
3.5. Involvement of reduced BDNF/p-mTOR in affective disorders of PPD-like mice
Activation of the BDNF/p-mTOR pathway may represent a unique mechanism for fast-acting antidepressants.35 Herein, the levels of BDNF and p-mTOR/mTOR were detected by Western blot in PPD-like mice. The results showed that the levels of BDNF were significantly decreased (F (3, 20) = 4.959, p < 0.01, Fig. 5A and B) in the hippocampus of PPD models, and PRE-084 or YL-0919 treatment restored BDNF to the normal level (PRE-084: p < 0.05; YL-0919: p < 0.05). Similarly, the p-mTOR levels among PPD-like mice were synergistically altered, but not mTOR (F (3, 20) = 5.592, p < 0.01, Fig. 5C). Moreover, coadministration with NE-100 reversed the benefit effect of YL-0919 in PPD-like mice (BDNF: F (2, 15) = 3.777, p < 0.05; p-mTOR: F (2, 15) = 4.335, p < 0.05).
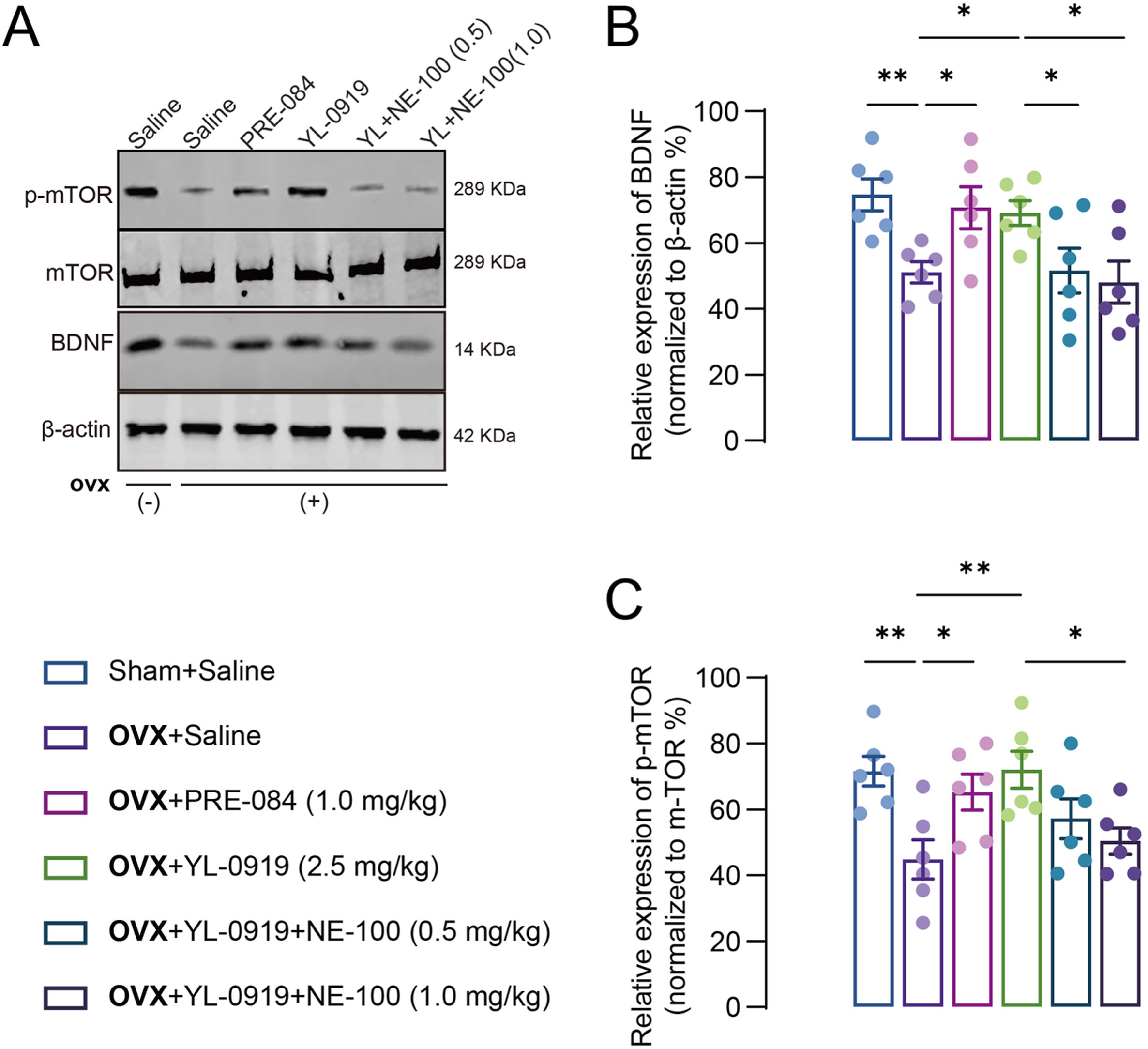
YL-0919 promoted the BDNF–mTOR pathway in PPD-like mice. Representative Western blot depicting the expression of BDNF, mTOR, and p-mTOR protein (A), in the hippocampus of PPD-like mice. Quantitative representation of BDNF, p-mTOR/mTOR protein expression in the hippocampus of PPD-like mice (B–C). Data are presented as mean ± SEM (n = 6 from three mice each group), *p < 0.05, **p < 0.01.
3.6. Effects of YL-0919 on neuroplasticity in the hippocampus of PPD-like mice
Defects in hippocampal neurogenesis are closely related to the pathogenesis of depression.36 We further assessed the effect of YL-0919 treatment on hippocampal neurogenesis in PPD-like mice using doublecortin (DCX) staining. We found a significant decrease in the number of DCX-positive immature neurons in the hippocampus of PPD-like mice compared with non-OVX-vehicle mice (F (3, 12) = 5.887, p < 0.01, Fig. 6A and B). YL-0919 or PRE-084 treated mice showed significantly increased DCX-positive cells in the dentate gyrus when compared to the OVX-vehicle mice (p < 0.05), and NE-100 treatment abolished the YL-0919 treatment effect on DCX-positive immature neurons (F (2, 9) = 4.025, p < 0.05). Moreover, the morphology of pyramidal neurons in the DG was observed by Golgi staining (Fig. 6C–E). The results revealed significant deficits in the total branching points of dendrites (F (3, 16) = 3.864, p < 0.05, Fig. 6C and D) and dendritic spine density (F (3, 16) = 6.970, p < 0.01, Fig. 6E and F) in PPD-like mice. Further drug treatment reversed the deficits in dendritic spine density in the DG of PPD-like mice. (YL-0919, p < 0.05; PRE-084, p < 0.01, Fig. 7D). These results indicate that the anxiolytic and antidepressant-like effect of YL-0919 in PPD-like mice are related to ameliorating the disruptions in dendritic complexity in the DG.
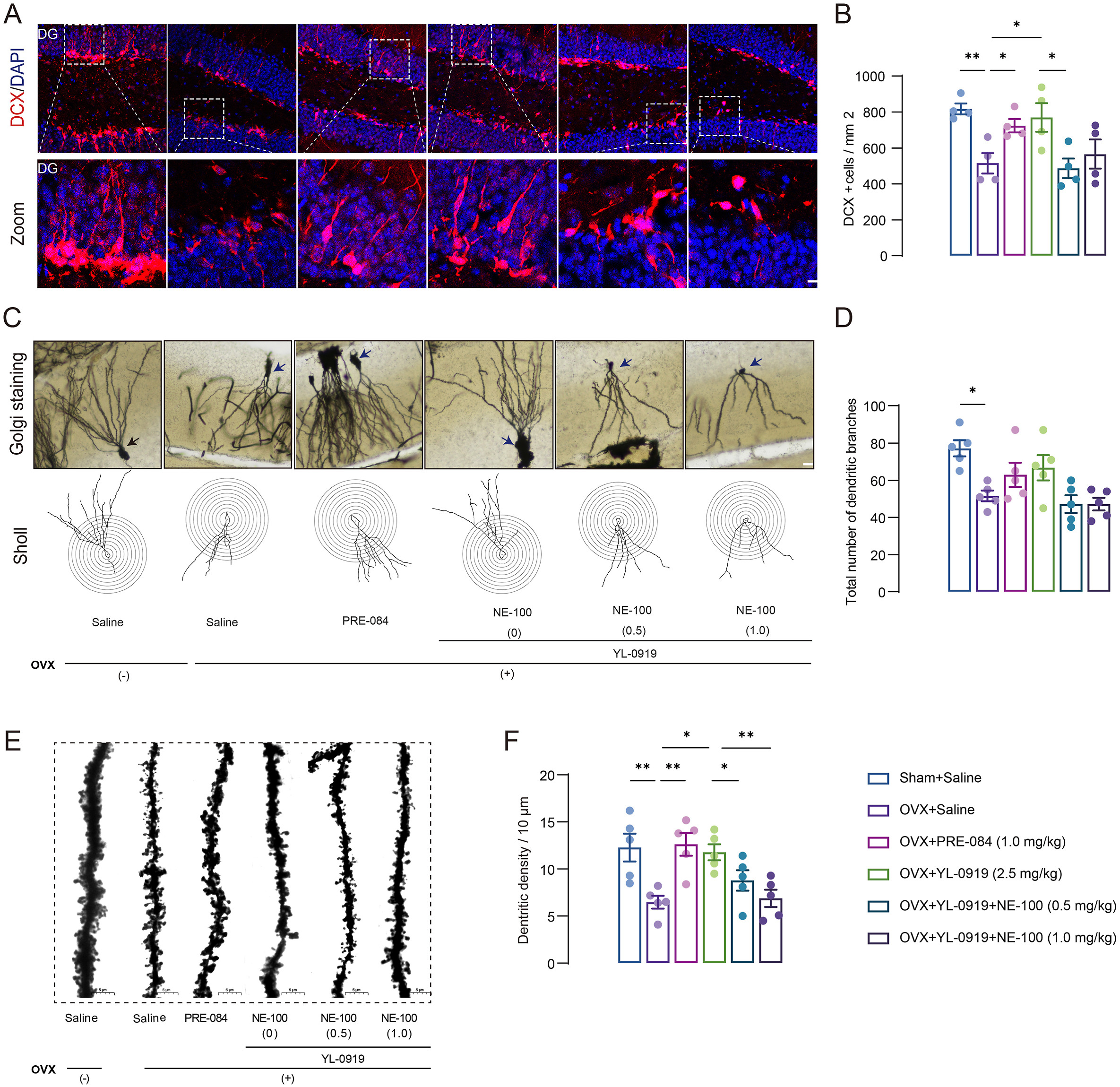
YL-0919 rescued hippocampal neuroplasticity deficiency in PPD-like mice.
(A–B) Representative immunostaining images of doublecortin (DCX) (red) and 4′,6-diamidi no-2-phenylindole (DAPI) (blue) in the dentate gyrus, Scale bar = 20 μm. YL-0919 and PRE-084 reversed the decrease in the number of DCX-positive cells in the dentate gyrus induced by estrogen withdrawal, and NE-100 blocked the effects of YL-0919. (C–D) Representative images of Golgi staining of pyramidal neurons in each group of DG areas and typical plots of sholl analysis of neuronal tracking, Scale bar = 20 μm. (E–F) Representative photomicrograph of dendritic spines from each group, Scale bar = 5 μm. Data are presented as mean ± SEM (n = 4–5/group), *p < 0.05, **p < 0.01. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
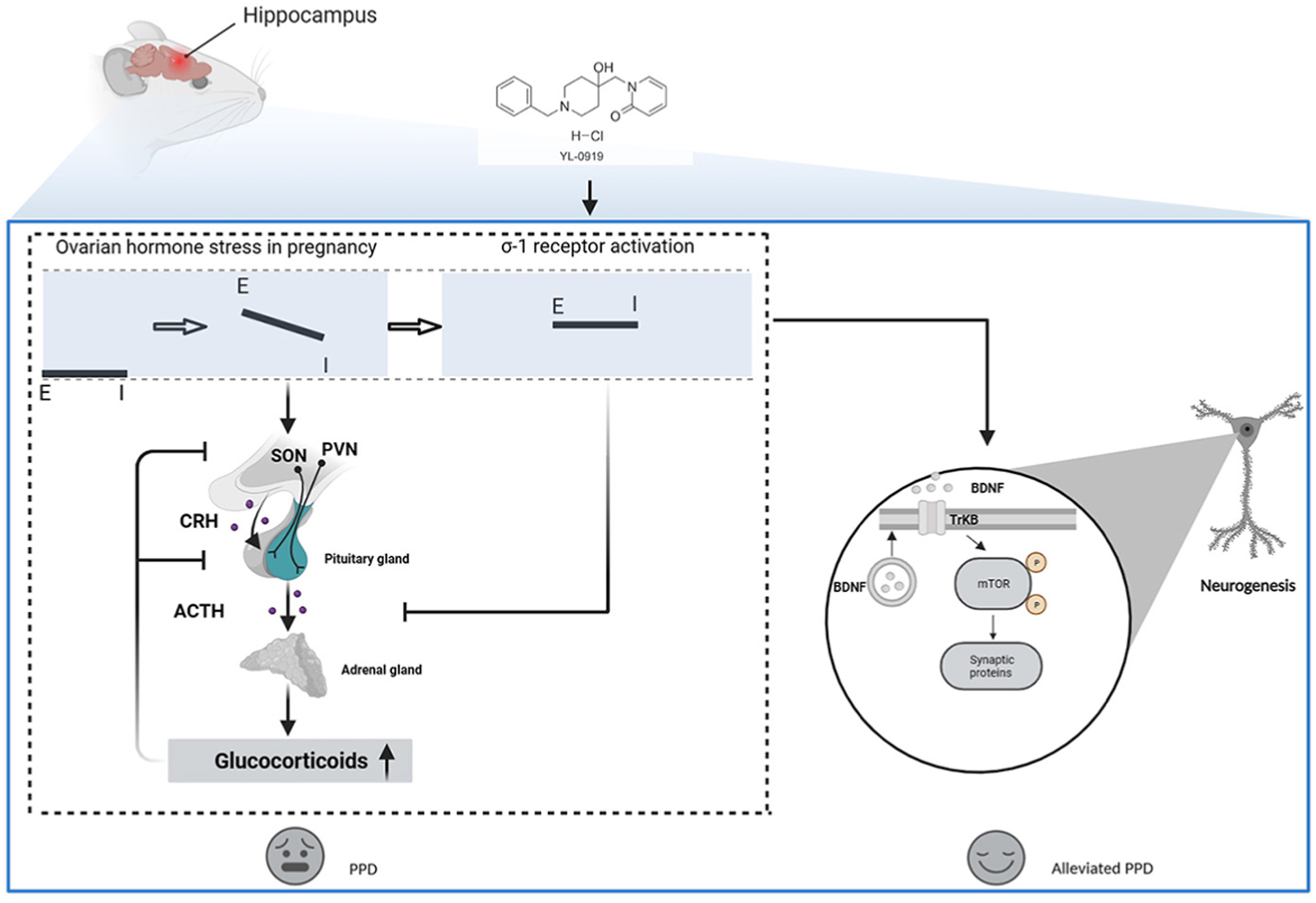
The potential mechanism action of YL-0919 in PPD.
The dramatic fluctuations in ovarian hormones during pregnancy and delivery can result in imbalances between GABAergic and glutamatergic signaling. In the PPD-like mice, the deficient GABAergic signaling is unable to suppress ovarian hormone stress-induced activation of the hypothalamic–pituitary–adrenal (HPA) axis, contributing to depression. Furthermore, σ-1 receptor agonist treatment offers the potential to accomplish a new balance in excitation/inhibition signal and restore the HPA axis. Ultimately, the re-established excitation/inhibition signal in the brain, including hippocampus causes the release of BDNF and improve neuro neuroplasticity in hippocampus, which may represent a unique mechanism for acting on antidepressants in PPD.
4. Discussion
PPD is disease proposed to been driven by multiple etiologies, including dramatic hormonal alterations during pregnancy, HPA axis activation, dysfunctional GABA/Glu signaling, and several others.37 In support of these, PPD-like behaviors can be elicited in a variety of rodent models.38 Among them, the “ovarian hormone withdrawal” hypothesis is the classical and widely used method to mimic the postpartum conditions, including abnormalities in monoamine neurotransmitters and receptors, metabolic products, and other biological factors, within the HPA axis, which cause the occurrence of PPD. In the present study, we employed this model not only to evaluate the effects of YL-0919 on PPD-like mice by multiple behavioral tests, but also to undermine the possible regulation mechanism of σ-1 receptor action in PPD. The results of this study report that the σ-1 receptor was activated by YL-0919 treatment, which accomplished GABA/Glu signaling rebalance and further restored the HPA axis signaling to the base level. Ultimately, the re-established GABA/Glu signaling in the brain causes release of BDNF, and increases neurogenesis in the hippocampus, which may represent a unique mechanism for the action of antidepressants in PPD.
The σ-1 receptor, designated an intracellular chaperone protein, is known to form direct interactions between several cellular proteins; it regulates a variety of cellular survival and metabolic functions, fine-tunes neuronal excitability, and modulates the transmission of information within brain circuits.39 In this study, we found that the σ-1 receptor is down-regulated in PPD-like mice, and the anxiety and depression-like behaviors of PPD-like mice were not only improved after drug treatment, but also the σ-1 receptor expression was restored to normal levels. A previous study in σ-1 receptor knockout (KO) animals also supports this; σ-1 receptor KO mice show depressive-like behaviors, particularly in female ovariectomized mice.40 These data imply that the σ-1 receptor play an important role in the etiology of PPD.
YL-0919, a novel structure-based compound developed in our laboratory and recently identified as a σ-1 receptor agonist, is also found to exhibit anxiolytic and anti-depressive-like behavior in PPD-like mice, which expands its pharmacological indications. The anxiolytic and antidepressant effects of YL-0919 in the PPD-like mice were blocked by NE-100, a potent selective σ-1 receptor antagonist. This is consistent with our previous results that the antidepressant effects of YL-0919 were totally reversed by another σ-1 receptor antagonist BD-1047.12These data provide further evidence that the σ-1 receptors are involved in the anxiolytic and antidepressant action of YL-0919. In addition, the current management strategy for PPD is similar with that used to treat major depressive disorder (MDD), with SSRIs being the commonly prescribed medications.37 However, antidepressant-based therapies can take weeks to months for a potential treatment effect to take hold.35 Our previous study in PPD-like rats demonstrated that sertraline did not show significant antidepressant effects until at least 3 weeks after a single gavage administration.2 In the present study, both agonists, YL-0919 and PRE-084, show significant anxiolytic and antidepressant effects after 1 week of drug administration, with a shorter onset of action compared to those of conventional antidepressants. Therefore, we believe that σ-1 receptor targeted antidepressants for PPD treatment may hold good promise.
It is well known that the medial prefrontal cortex (mPFC) or hippocampus relies on the dynamic balance of excitation/inhibition neurotransmitters for the fundamentals of normal neurological functions.41,42 Glu is the main excitatory neurotransmitter in the mammalian brain, and GABA is the main inhibitory neurotransmitter. Multiple lines of evidence also further support the link between GABAergic signaling with PPD.37 In animal models, fluctuations in the expression and function of GABAA receptor subunits have been observed during pregnancy and the postpartum period.5 Little evidence in the published reports is available in relation to the changes in the Glu system during the postpartum period. A novel and significant finding of the present study is that the enhanced glutamatergic signaling and decreased GABAergic signaling in hippocampus was intimately associated with increased anxiety and depressive stress responses in PPD-like mice, which is in line with a previous clinical study showing an upregulation Glu levels in women with PPD compared to healthy controls.43 Further results in our study show that treatment of PPD-like mice with an σ-1 receptor agonist restored GABAergic signaling to normal levels, accompanied by alleviated anxiety and depressive-like behavior, whereas glutamatergic signaling has the opposite action. In addition, we found that σ-1 agonists can affect excitation/inhibition signal in specific brain regions by modulating the release of gliotransmitters to ameliorate anxiety-depression-like behaviors. Therefore, we speculate that the ovarian hormonal stress during pregnancy induces excessive glutamatergic signaling, resulting in multiple defects in GABAergic transmission and exacerbating stress-induced glutamatergic hyperexcitability, resulting in failure to inhibit activation of the stress HPA axis (Fig. 7). In support of this hypothesis, hyperactivity of the HPA axis (elevated levels of ACTH and CORT in the serum) was observed in PPD-like mice in our study, while σ-1 agonist YL-0919 treatment force ACTH and CORT to return to baseline level. Collectively, these points of evidence reinforce the hypothesis that there is a link between GABAergic signaling and the HPA axis, although we need more studies to demonstrate whether activation of the σ-1 receptor regulates this link directly or indirectly. Current findings have changed our conceptualization of the pathophysiology and treatment of postpartum depression and have encouraged the development of depression treatments based on σ-1 receptor research.
A recent study has reported that the BDNF–mTOR pathway in the nucleus accumbens were downregulated in post-traumatic stress disorder rat models, whereas this downregulation was recovered by a metabolite of ketamine, (2R, 6R)-HNK, administration.44 Accordingly, we found that the BDNF–mTOR pathway was deficient in the hippocampus, whereas YL-0919 treatment restores this defect along with alleviated anxiety–depressive-like behavior in PPD-like mice, thus normal functioning of the BDNF–mTOR pathway is essential for σ-1 receptor agonists to exert their anxiolytic and antidepressant effects. mTOR is a downstream modulator of BDNF that plays a key role in brain-specific processes, such as learning and synaptic plasticity.44 Herein, DCX immunofluorescence and Golgi staining also provided similar evidence that YL-0919 can promote hippocampal neurogenesis in PPD-like mice. Taken together, the present study suggests that the beneficial effects of YL-0919 on mood and behavior in PPD-like mice are associated with in vivo σ-1 receptor activation and improved neurogenesis to enhance synaptic plasticity (Fig. 7).
Given these results and the previous work described above, σ-1 receptors are highly likely to be involved in this complex regulatory mechanism in PPD. Nevertheless, there are several limitations in the present research. On the one hand, we observed that σ-1 receptors are involved in the modulation of the GABA/Glu signaling system in PPD; however, data obtained with different methods cannot be integrated into a single coherent picture, and multidisciplinary approaches will be required to grasp the significance between the σ-1 receptor and GABA/Glu system in maintaining normal excitation/inhibition signal. On the other hand, the current study mainly focuses on the hippocampus brain region; other regions may also be responsible for the progression of PPD. Thus, it would be interesting to observe region-specific regulation of BDNF–mTOR signaling by the activation of the σ-1 receptor, such as the dorsal and ventral hippocampus, mPFC, and amygdala.
In summary, these findings highlight how YL-0919 can exert widespread effects on the hippocampus region of the brain, providing new insight into neuron-specific synaptic plasticity mechanisms. YL-0919 rapidly restore the excitation/inhibition signal in the brain by activating σ-1 receptors to modulate GABA/Glu signaling, and then normalize the HPA axis signal, thus promoting hippocampal neuroplasticity. Novel compounds targeting the σ-1 receptor may inform the advancement of novel treatments for PPD.