
Transit and integration of extracellular mitochondria in human heart cells

By Douglas B. Cowan, Rouan Yao, Jerusha K. Thedsanamoorthy, David Zurakowski, Pedro J. del Nido, and James D. McCully
Excerpt from the article published in Scientific Reports 7, 17450 (2017). Published: , DOI: https://doi.org/10.1038/s41598-017-17813-0
Editor’s Highlights
- Mitochondria play an essential role in energy production and cellular homeostasis.
- Dysfunction of these organelles as a result of ischemia or genetic mutations can lead to the loss of high-energy phosphate reserves, accumulation of mitochondrial calcium, and a buildup of reactive oxygen molecules.
- Transplantation of isolated mitochondria to the ischemic heart leads to reductions in infarct size, increases in adenosine triphosphate (ATP) production, and improvements in contractility.
- The uptake and subsequent fusion of extracellular mitochondria with recipient cell mitochondria is an evolutionarily-conserved and pervasive biological process.
- Several proteins are involved in fusion of exogenous mitochondria with endogenous mitochondria.
- A complete understanding of the mechanisms that bring about endocytosis of extracellular mitochondria and fusion of those organelles with the endogenous mitochondrial network may provide a foundation for innovative new treatments to a number of human diseases and disorders.
Abstract
Tissue ischemia adversely affects the function of mitochondria, which results in impairment of oxidative phosphorylation and compromised recovery of the affected organ. The impact of ischemia on mitochondrial function has been extensively studied in the heart because of the morbidity and mortality associated with injury to this organ. As conventional methods to preserve cardiac cell viability and contractile function following ischemia are limited in their efficacy, we developed a unique approach to protect the heart by transplanting respiration-competent mitochondria to the injured region. Our previous animal experiments showed that transplantation of isolated mitochondria to ischemic heart tissue leads to decreases in cell death, increases in energy production, and improvements in contractile function. We also discovered that exogenously-derived mitochondria injected or perfused into ischemic hearts were rapidly internalised by cardiac cells. Here, we used three-dimensional super-resolution microscopy and transmission electron microscopy to determine the intracellular fate of endocytosed exogenous mitochondria in human iPS-derived cardiomyocytes and primary cardiac fibroblasts. We found isolated mitochondria are incorporated into cardiac cells within minutes and then transported to endosomes and lysosomes. The majority of exogenous mitochondria escape from these compartments and fuse with the endogenous mitochondrial network, while some of these organelles are degraded through hydrolysis.
Introduction
Mitochondria play an essential role in energy production and cellular homeostasis. Dysfunction of these organelles as a result of ischemia or genetic mutations can lead to the loss of high-energy phosphate reserves, accumulation of mitochondrial calcium, and a buildup of reactive oxygen molecules1,2,3,4,5. Our previous studies demonstrated that transplantation of isolated mitochondria to the ischemic heart leads to reductions in infarct size, increases in adenosine triphosphate (ATP) production, and improvements in contractility6,7. We also observed that mitochondria injected or perfused into the heart were rapidly internalised by a variety of cardiac cells including cardiomyocytes and fibroblasts7,8. Additional experiments using cell cultures proved that the uptake of mitochondria occurs through actin-dependent endocytosis and results in rescue of cellular function by increasing energy production and replenishing mitochondrial DNA (mtDNA)9. Although other researchers have observed endocytic incorporation of extracellular mitochondria, the intracellular trafficking and fate of these organelles remains unknown10,11,12,13,14,15.
In this study, we used three-dimensional super-resolution structured illumination microscopy (3-D SR-SIM) and transmission electron microscopy (TEM) to reveal the intracellular position of endocytosed mitochondria in human induced pluripotent stem cell-derived cardiomyocytes (iPS-CMs) and human cardiac fibroblasts (HCFs). By labelling isolated mitochondria with fluorescent proteins or gold nanoparticles, we were able to observe the transit of exogenous mitochondria in these cells. Distinct fluorescent labelling of various cell compartments in iPS-CMs and HCFs allowed us to visualise the progression of exogenous mitochondria through the endolysosomal system and establish that these organelles primarily integrate with the endogenous mitochondrial network in both cardiac cell types. Immunoblot experiments confirmed that the cardiomyocytes and fibroblasts used in these studies expressed proteins compatible with mitochondrial fusion.
When combined with the findings of other investigators, our results strongly support the notion that the uptake and subsequent fusion of extracellular mitochondria with recipient cell mitochondria is an evolutionarily-conserved and pervasive biological process7,8,9,10,11,12,13,14,15,16. A thorough understanding of the endocytic uptake, intracellular transit, and mitochondrial integration of exogenous mitochondria in cells may present new treatment strategies for the ischemic heart and drive the development of organelle-based therapeutics for a host of other human diseases and disorders17,18,19,20.
Results
Labelling of organelles and characterisation of isolated mitochondria
We investigated the temporal and spatial fate of endocytosed mitochondria in non-dividing iPS-CMs and dividing HCFs. The identity and morphology of these cardiac cells was substantiated by immunostaining with α-actinin (ACTN) and vimentin and both cell types were shown to react well with established mitochondrial antibodies (TOMM20 or MTC02) (Extended Data Fig. 1a). To discern exogenous mitochondria within cultured cells, we labelled HCF mitochondria with green fluorescent protein (GFP) and used red fluorescent proteins (RFP) to label various HCF and iPS-CM cell compartments through baculovirus-mediated transfer of mammalian fusion genes (Fig. 1a). Both cell types were readily infected with baculoviruses carrying fluorescent protein genes and exhibited specific expression of GFP or RFP in organelles including mitochondria, early and late endosomes, lysosomes, Golgi complexes, and the endoplasmic reticulum (Extended Data Fig. 1b). Isolated HCF GFP-labelled mitochondria were stained with MitoTracker Red CMXRos or a human mitochondria-specific antibody (MTC02) to confirm their identity and then imaged using 3-D SR-SIM (Fig. 1b). Isolated mitochondria were generally spherical in shape and varied in diameter from 250 to 2000 nm with the majority of these organelles falling within the 350 to 600 nm range7. In addition, adenosine triphosphate (ATP) measurements verified that isolated mitochondria were viable and energised (Fig. 1c), while transmission electron microscopy revealed these organelles were structurally intact (Fig. 1d)7,8,21.
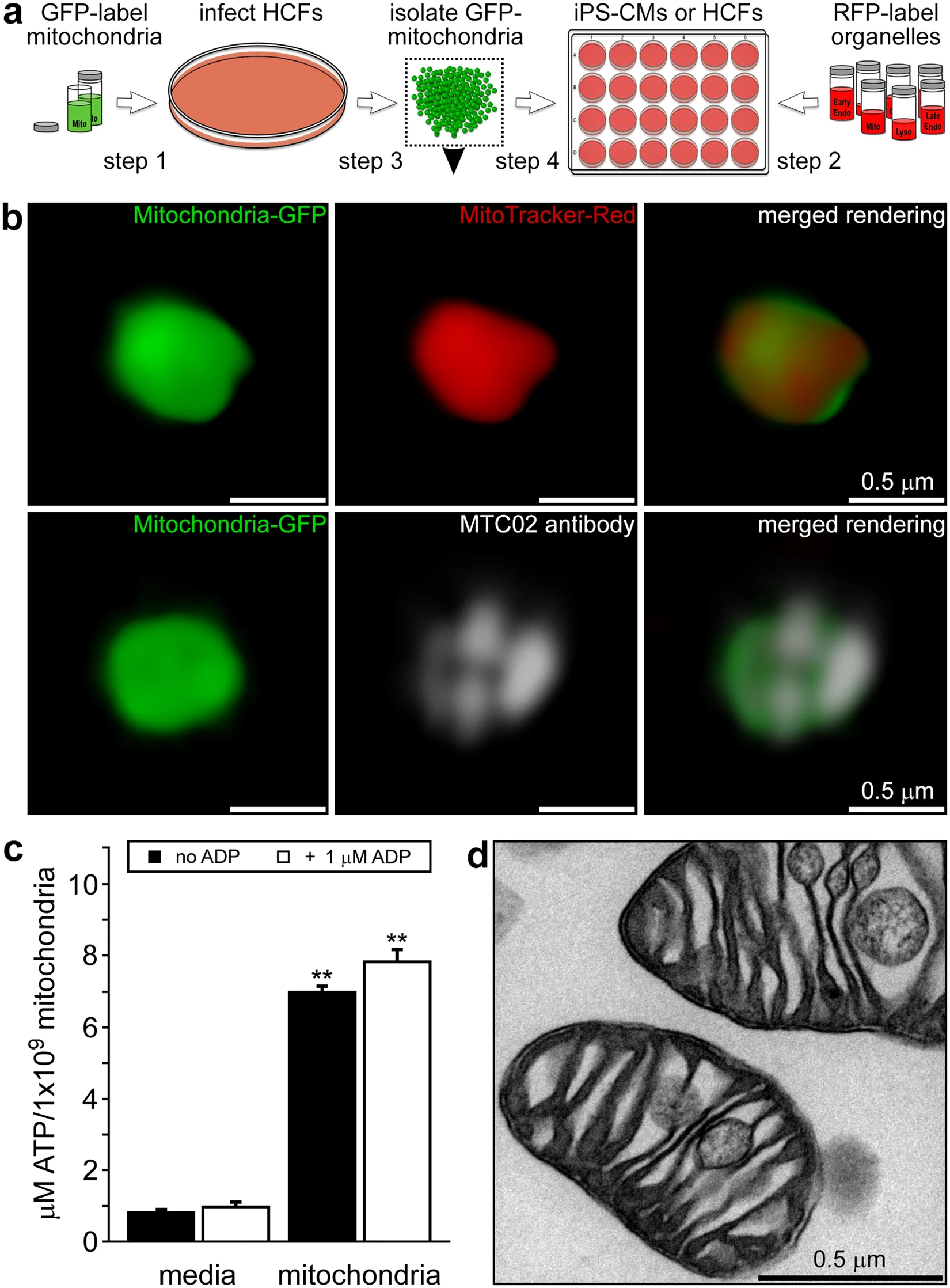
Experimental strategy and characterisation of isolated human fibroblast mitochondria. (a) HCFs infected with BacMam CellLight Mitochondria-GFP were used for mitochondrial isolations and iPS-CMs or HCFs on coverslips were infected with RFP CellLight reagents to label specific cell structures. Isolated GFP-labelled mitochondria were added to RFP-labelled cells for 0 to 4 h. (b) 3-D SR-SIM of isolated HCF mitochondria stained with MitoTracker Red CMXRos (top panels) or the human mitochondria-specific antibody MTC02 (bottom panels). Isolated mitochondria, stained mitochondria, and the combined image are shown left to right. Scale bars, 0.5 µm. (c) ATP content in media or isolated HCF mitochondria in the presence or absence of 1 µM ADP. Error bars indicate standard error of the mean. ATP levels were significantly higher in mitochondria than in media-only control groups (Student’s t-test, **p < 0.001). (d) Transmission electron microscopy of isolated HCF mitochondria. Scale bar, 0.5 µm. The image is representative of four separate mitochondrial isolations.
Endocytosis and intracellular position of exogenous mitochondria
To examine endocytosis of isolated mitochondria in iPS-CMs, we first labeled HCF mitochondria with 10 nm diameter gold nanoparticles (Fig. 2a) to allow for their identification by transmission electron microscopy. Labelled exogenous mitochondria were observed outside cells, near cell surfaces, in the process of endocytic engulfment, and inside iPS-CMs22. Next, the intracellular position of endocytosed mitochondria in iPS-CMs was determined using four color 3-D SR-SIM (Fig. 2 and Extended Data Fig. 2). Because we did not observe GFP-labelled mitochondria associated with either the Golgi complex or endoplasmic reticulum, we focused on examining whether RFP-labelled endosomes, lysosomes, or endogenous mitochondria colocalised with internalised HCF mitochondria. Early endosomes (Fig. 2b), late endosomes (Fig. 2c), and lysosomes (Fig. 2d) of cardiomyocytes were all found to contain GFP-labelled mitochondria at each time examined (0.5, 1, 2, and 4 h). In addition, endocytosed mitochondria were found in close proximity to the endogenous mitochondrial network, which was generally located apical to the contractile apparatus (Extended Data Fig. 2a and Movie S1).
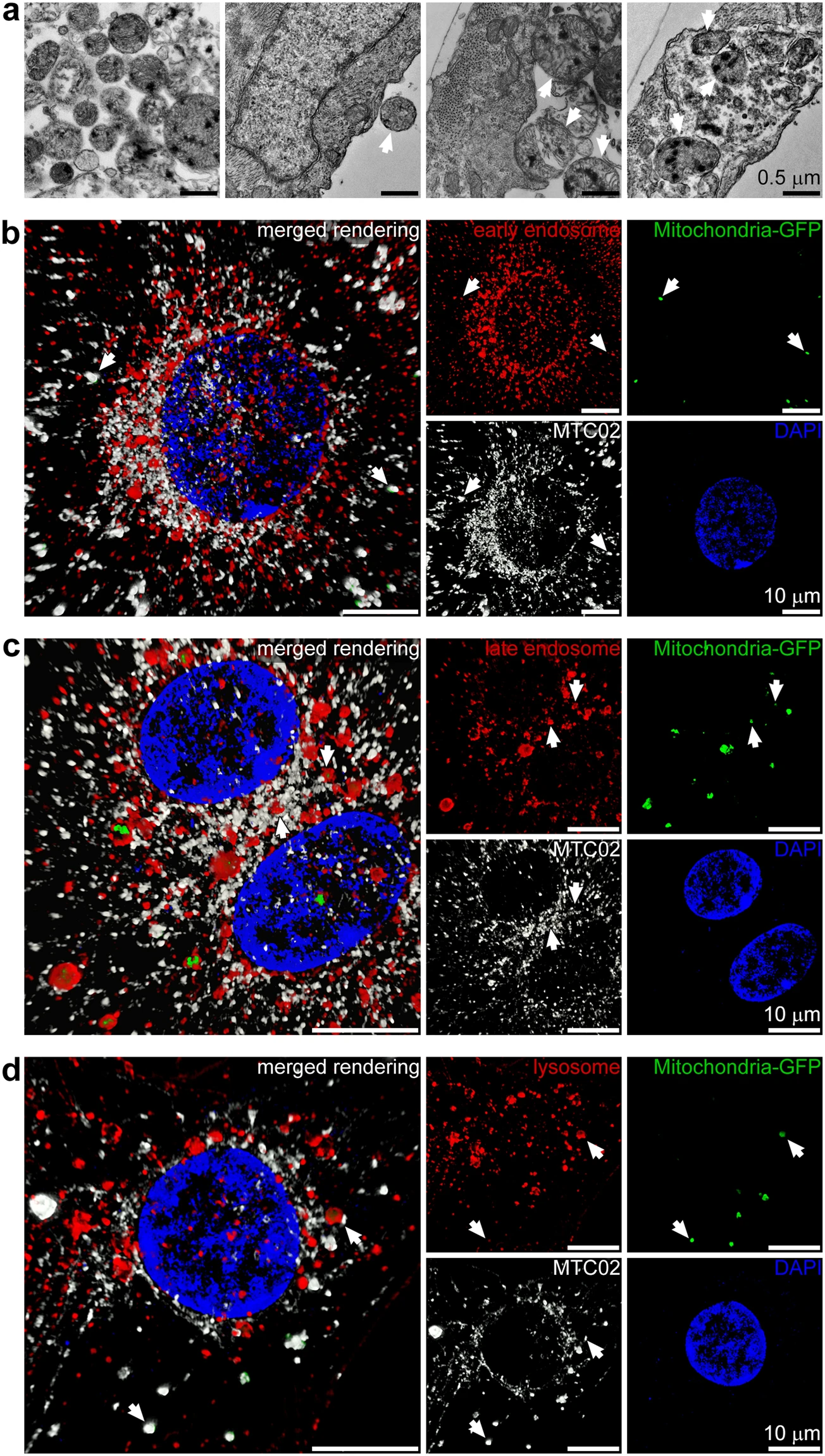
Uptake and endocytic transport of extracellular mitochondria in cardiomyocytes. (a) iPS-CMs were exposed to gold-labelled HCF mitochondria for 2.5, 5, and 10 min and imaged by TEM. Labelled mitochondria had electron dense deposits and were apparent at all three study times outside cells, adjacent to the apical cell surface, undergoing endocytosis, and inside cells (left to right and indicated by arrows). Images are representative of four experiments at 5 min. Scale bars, 0.5 µm. (b) Four color 3-D SR-SIM of colocalisation of exogenous mitochondria (green) with early endosomes (red) at 4 h. Nuclei were stained with DAPI (blue) and both endogenous and exogenous mitochondria were detected with the MTC02 antibody (white). The combined image (left) and each color (right) are shown. Scale bars, 10 µm. (c) Colocalisation of HCF mitochondria (green) with late endosomes (red) at 1 h. Scale bars, 10 µm. (d)Colocalisation of mitochondria (green) with lysosomes (red) at 4 h. Scale bars, 10 µm. In (c,d) arrows indicate examples of exogenous mitochondria associated with each compartment, which also reacted with the anti-mitochondria MTC02 antibody. Nuclei were stained with DAPI (blue) and images represent twelve separate volumes analysed at each time (0.5, 1, 2, and 4 h).
Escape of exogenous mitochondria from the endolysosomal system
Exogenous mitochondria associated with early endosomes, late endosomes, and lysosomes were observed to be fully encapsulated or partly contained within these compartments. Mitochondria partially enclosed in endocytic vesicles were considered to be escaping from their respective endosomal or lysosomal compartments. Cell preparations were stained with anti-human mitochondria antibody (MTC02) to confirm that green fluorescence was attributable to the presence of intact organelles; though, this antibody was less reactive with encapsulated mitochondria—possibly a consequence of reduced antigen accessibility. To further corroborate the identity of the above compartments, we stained cardiomyocyte cell preparations with early endosome antigen 1 (EEA1) and the late endocytic marker Rab7, which helps control aggregation and fusion of late endosomes and lysosomes (Extended Data Fig. 2b–d)23. In addition, we observed some lysosomes associated with exogenous mitochondria stained for Rab7, which could simply epitomise the heterogeneity of the lysosomal compartment or indicate these particular compartments fused into endolysosomal vesicles24,25. We also noticed exogenous mitochondria in lysosomes that were Rab7 negative; however, we did not quantitate the proportion of internalised mitochondria in specific endocytic compartments expressing distinct protein compositions.
The escape of exogenous mitochondria from endosomes and lysosomes was assessed by counting colocalisation of GFP-labelled mitochondria with each of these compartments. High-magnification 3-D SR-SIM suggested GFP-labelled mitochondria became increasingly reactive with MTC02 antibody upon emerging from late endosomes (Fig. 3a and Movie S2) as well as early endosomes and lysosomes (not shown). Enumeration of colocalisation of endocytosed mitochondria with endosomes and lysosomes at each time established that a majority of these exogenous organelles were associated with these vesicles at the earlier times (0.5 to 2 h); however, there was no obvious pattern to endolysosomal trafficking other than a decline in colocalisation at 4 h (Fig. 3b). The number of mitochondria escaping from these compartments showed the opposite trend with the largest number of organelles emerging from late endosomes and lysosomes at 4 h (Fig. 3c). We extended these analyses to include colocalisation between endogenous and exogenous mitochondria (Fig. 3d). These experiments demonstrated a large percentage of internalised GFP-labelled mitochondria were closely associated with the cardiomyocyte mitochondrial network from 0.5 to 4 h. The functional consequence of the uptake and incorporation of extracellular mitochondria in cardiomyocytes was a sustained and significant increase in ATP production, similar to our earlier findings9. Together, these results signify that endocytosis and intracellular transportation of extracellular mitochondria in cardiomyocytes is an ongoing and efficient biological process.
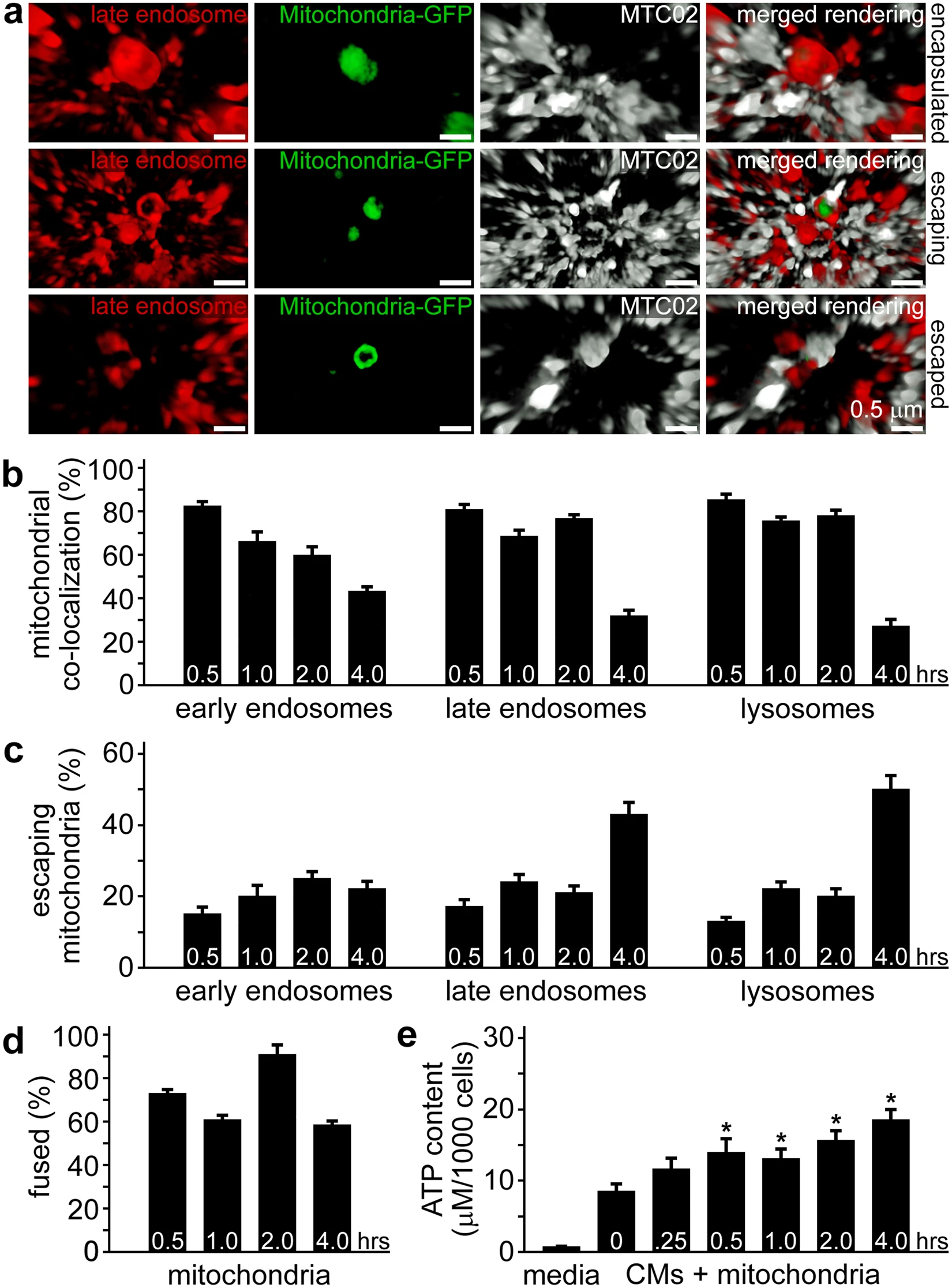
Encapsulation and escape of internalised mitochondria from cardiomyocyte cell compartments. (a) Late endosomes (red) associated with GFP-labelled exogenous mitochondria (green) were imaged by 3-D SR-SIM. Examples of encapsulated, escaping, and escaped mitochondria are shown after 1 h of incubation of isolated mitochondria and iPS-CMs. The anti-mitochondria antibody MTC02 demonstrated increased reactivity with exogenous mitochondria as they emerged from late endosomes and escaped mitochondria (bottom panels) become completely enveloped by MTC02. Images are representative of four experiments. Scale bars, 0.5 µm. (b) Quantitation of mitochondrial colocalisation with early endosomes, late endosomes, and lysosomes at 0.5, 1, 2, and 4 h. Results are expressed as the percentage of mitochondria associated with each compartment (i.e. encapsulated). (c) Quantitation of mitochondria escaping early endosomes, late endosomes, and lysosomes at 0.5, 1, 2, and 4 h. Results are expressed as the percentage of mitochondria associated with each compartment, but not fully encapsulated and represent a minimum of twelve experiments. (d) Quantitation of the colocalisation of endocytosed mitochondria with endogenous cardiomyocyte mitochondria. Results are expressed as the percentage of GFP-labelled mitochondria closely associated with RFP-labelled mitochondria. (e) ATP content of culture media and cardiomyocytes treated with isolated HCF mitochondria for 0, 0.25, 0.5, 1, 2, and 4 h. ATP content is expressed as µM per 1000 cells and error bars indicate standard error of the mean. ATP was significantly higher in iPS-CMs treated with mitochondria for 0.5, 1, 2, and 4 h compared to 0 h untreated cells (Student’s t-test, *p < 0.02). Results were obtained from twelve separate experiments.
Fusion of exogenous mitochondria with the endogenous mitochondrial network
The fusion of endocytosed mitochondria with the endogenous mitochondria of cardiomyocytes (Fig. 4a and Movie S3) and cardiac fibroblasts (Extended Data Fig. 3) was assessed by 3-D SR-SIM. In combination with depth coding analyses (Extended Data Fig. 4aand Extended Data Fig. 3a), these experiments demonstrated that exogenous mitochondria fused with the endogenous mitochondrial network after escaping from endolysosomal compartments at 0.5, 1, 2, and 4 h. The presence of mitochondrial fusion-associated proteins was assessed by studying the expression of mitofusin-1 (MFN1), mitofusin-2 (MFN2), and optic atrophy 1 (OPA1) proteins in iPS-CMs and HCFs (Fig. 4b). Whole cell and isolated mitochondrial lysates were analysed for fusion proteins by immunoblot analyses. The expression and phosphorylation of the mitochondrial fission-associated dynamin-related protein 1 (DRP1) and the presence of the mitophagy-associated protein Parkin (Extended Data Fig. 4b) were determined for comparison. The anti-mitochondria antibodies MTC02 and 113-1 were used to establish equivalent loading of lysates in polyacrylamide gels8. We subsequently examined the spatial distribution of mitofusin proteins in cardiomyocytes using 3-D SR-SIM (Fig. 4c, Extended Data Fig. 4c, and Movie S4).
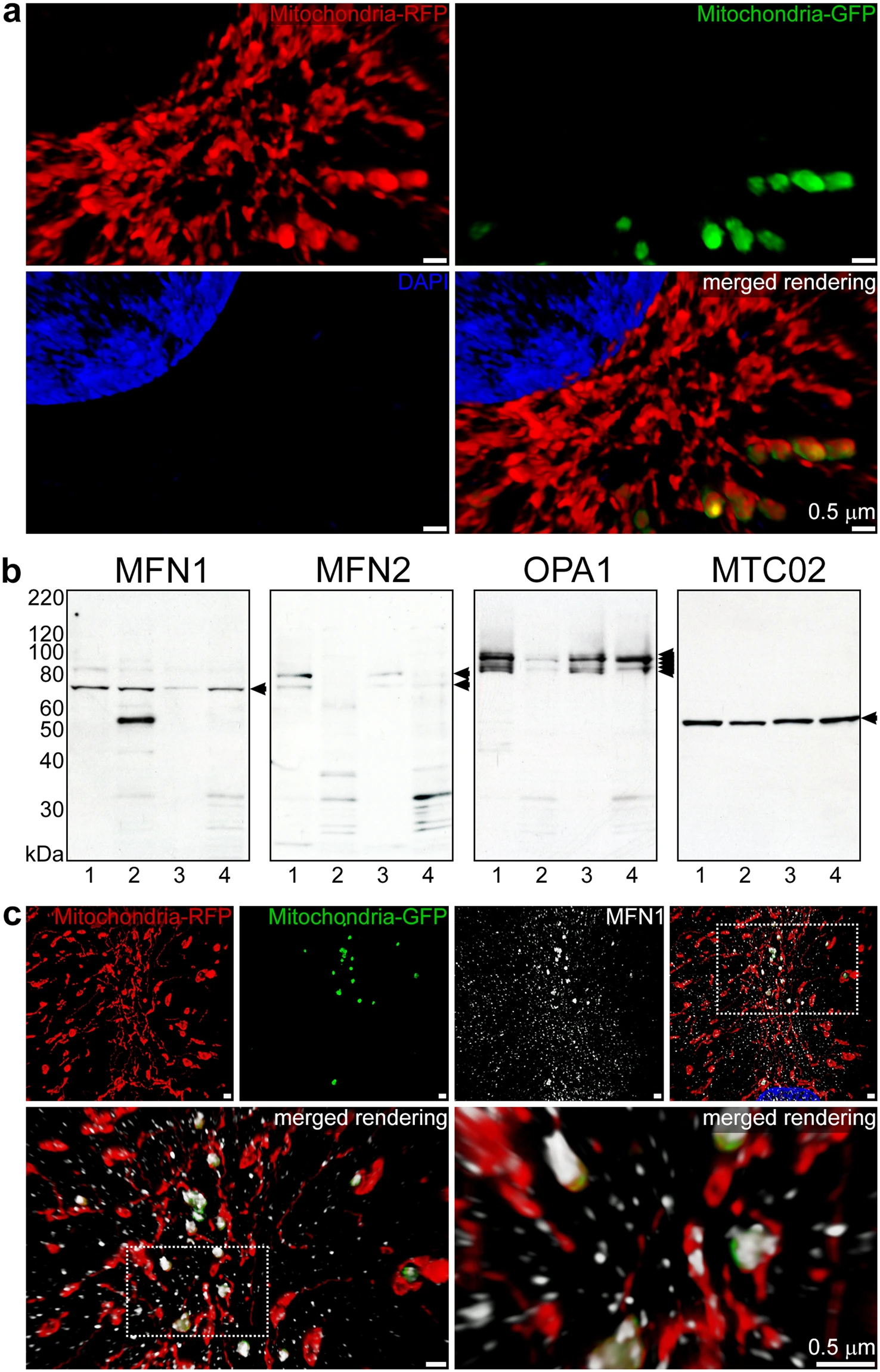
Fusion of exogenous mitochondria with the cardiomyocyte mitochondrial network. (a) iPS-CMs with RFP-labelled mitochondria (red) treated for 4 h with isolated GFP-labelled HCF mitochondria (green). After fixation, the nuclei were stained with DAPI (blue) and slides were imaged using 3-D SR-SIM. Individual red, green, and blue channels are shown along with the combined color rendering. Fusion of endocytosed mitochondria with the endogenous mitochondrial network is readily apparent. Mitochondrial fusion was apparent at all times examined (0.5, 1, 2, and 4 h) and four separate experiments were performed. Scale bars, 0.5 µm. (b) Immunoblot analysis of whole cell and isolated mitochondrial lysates (25 µg per lane) from iPS-CMs and HCFs using antibodies directed against MFN1, MFN2, OPA1, and mitochondria (MTC02). Lanes 1 to 4 contained the following lysates: iPS-CM cell proteins, HCF cell proteins, iPS-CM mitochondrial proteins, and HCF mitochondrial proteins. Arrows indicate the specifically detected protein(s) for each antibody and immunoblot experiments were repeated six times. (c) 3-D SR-SIM of human cardiomyocytes containing RFP-labelled mitochondria (red) incubated with isolated GFP-labelled exogenous mitochondria (green) for 30 min. Coverslips were fixed and stained with a mitofusin-1 (MFN1) antibody (white). The top panels show the three separate color channels and the combined image (left to right). The boxed regions have been magnified in successive images (top right to lower panels, left to right) to show the reactivity of the MFN1 antibody with fusing mitochondria and the presence of this antibody throughout the cytosol. Parallel experiments were performed at 1, 2, and 4 h and the presented images are representative of four separate experiments. Scale bars, 0.5 µm.
Whole cell lysates from iPS-CMs and HCFs contained equivalent amounts of MFN1 protein (Fig. 4b, lanes 1 and 2); however, only cardiomyocytes had detectable quantities of MFN2 (Fig. 4b, lane 1). In contrast, examination of lysates from isolated mitochondria revealed cardiomyocytes had relatively less MFN1 associated with these organelles (Fig. 4b, lane 3). HCFs had nearly equal levels of this protein in whole cell and mitochondrial lysates (Fig. 4b, lanes 2 and 4). It is worth noting that the MFN1 antibody strongly reacted with an approximately 55 kDa protein in HCF whole cell lysates. This could indicate the presence of a protein related to MFN1 in HCFs that was not associated with mitochondria. Isolated mitochondria from both cell types contained low levels of MFN2 (Fig. 4b, lanes 3 and 4). Analysis of OPA1 expression showed cardiomyocytes possessed relatively large quantities of long and short isoforms of this protein, whereas HCFs produced fewer variants in reduced quantities (Fig. 4b, lanes 1 and 3 versus 2 and 4). A comparison of whole cell lysates with mitochondrial lysates confirmed OPA1 was largely associated with mitochondria (Fig. 4b, lanes 1 and 2 versus lanes 3 and 4). Identical immunoblots confirmed equal reactivity of each lysate with MTC02 (Fig. 4b) and 113–1 (Extended Data Fig. 4b). DRP1 and a phosphorylated variant of DRP1 (S616) were expressed in both iPS-CMs and HCFs (Extended Data Fig. 4b, lanes 1 and 2). Evaluation of mitochondrial lysates revealed a small amount of DRP1 protein in iPS-CMs (Extended Data Fig. 4b, lane 3); but, S616 phosphorylated DRP1 was not observed (Extended Data Fig. 4b, lane 3). Whole cell and mitochondrial lysates from fibroblasts contained nearly equal levels of DRP1 (Extended Data Fig. 4b, lane 2 and 4) and DRP1 S616 (Extended Data Fig. 4b, lanes 2 and 4). Additional immunoblot analysis of DRP1 S637 phosphorylation using the same protein lysates failed to detect this post-translational modification (not shown); however, Parkin was present in whole cell lysates from HCFs (Extended Data Fig. 4b, lane 2).
Imaging of the spatial distribution of MFN1 and MFN2 using 3-D SR-SIM demonstrated that these proteins localised to fusing exogenous and endogenous mitochondria in addition to a multitude of other intracellular sites (Fig. 4c and Extended Data Fig. 4c). Even though MFN1 and MFN2 were observed to be concentrated at the sites of fusion (Fig. 4c, Movie S4, and Extended Data Fig. 4c), both mitofusin proteins were widely dispersed throughout the cytosol and nuclei of cardiomyocytes.
Together, our results prove that extracellular mitochondria are rapidly and efficiently endocytosed in human iPS-CMs and primary HCFs and the majority of these exogenous organelles fuse with the endogenous mitochondrial network (Extended Data Fig. 5). Cardiomyocyte and fibroblast mitochondria contain fusion-associated proteins (e.g. MFN1, MFN2, and OPA1), which probably facilitates this process. At the same time, some exogenous mitochondria do not appear to escape from endolysosomal compartments and presumably undergo hydrolysis and degradation through autophagy—an assumption supported by the appearance of fragmented mitochondria in some lysosomes (Extended Data Fig. 2d).
Discussion
The ability to distinguish the precise intracellular location of internalised exogenous human mitochondria using 3-D super-resolution microscopy and transmission electron microscopy in cardiac cells has allowed us to understand the intracellular trafficking and fate of these organelles through time. Electron microscopy revealed that extracellular mitochondria begin to be endocytosed by human iPS-derived cardiomyocytes (iPS-CMs) and primary human cardiac fibroblasts (HCFs) within minutes. These exogenous organelles successively progress through the endolysosomal system from early endosomes to late endosomes to lysosomes. While some of these mitochondria appear to be destined for degradation, our results provide compelling evidence that the majority of exogenous mitochondria escape from endosomal and lysosomal compartments and effectively fuse with endogenous cardiac cell mitochondria. Our findings in iPS-CMs and HCFs support the idea that endocytosis and integration of exogenous mitochondria is an evolutionarily-conserved biological process that is distinct from recently described cell-to-cell transfer of mitochondria through tunneling nanotubes or exosomes16,26,27,28. In essence, our observations of the uptake of isolated mitochondria in human heart cells corroborates the endosymbiotic theory advanced and substantiated by Lynn Margulis (née Sagan) half a century ago29.
Our earlier experiments, and those of others, established that endocytosis of extracellular mitochondria was dependent on actin polymerisation9,11,14 and that internalisation of these organelles resulted in long-term replenishment of mtDNA in HeLa cell-derived ρ0 cells9,28,30, which are devoid of mtDNA31. Restoration of the ρ0 mitochondrial genome through the uptake of mitochondria isolated from HeLa cells led to enhancement of intracellular ATP levels and oxygen consumption rates. Because increases in ρ0 cellular respiration persisted for more than 50 population doublings, it was evident that endocytosed HeLa cell mitochondria containing functional mtDNA were lastingly integrated within these cells9. In spite of this discovery, we did not resolve how internalised HeLa cell mitochondria were transported within ρ0 cells or determine whether they fused with endogenous mitochondria, persisted as distinct organelles, or merely contributed their DNA to ρ0 mitochondria. The same can be said for our previous experiments, which established internalisation of extracellular mitochondria in cardiac cells7,8.
In this study, we defined the intracellular transit of extracellular mitochondria from the plasma membrane through to their incorporation with the endogenous mitochondrial network in two types of human heart cells—dividing HCFs and non-dividing iPS-CMs. Purified iPS-CMs represent a suitable experimental model as these spontaneously-beating cells do not divide and retain the biochemical and electrophysiological responses typical of human cardiac myocytes. We anticipated that these terminally differentiated cells would more accurately predict in vivo responses compared to other culture models like immortalised cells or cardiac progenitors, which exhibit a less definitive differentiation potential32. Because iPS-CMs were reprogrammed using non-integrating episomal vectors and cultured without feeder cells, they had a consistent phenotype with no pathogenic contaminants. At the same time, our cardiomyocyte cultures displayed a uniform ‘late phase’ cardiomyocyte phenotype with a well-developed mitochondrial network and organized contractile apparatus33. For comparison, we examined primary HCFs, which were not derived from iPS cells7. We found that both cell types could efficiently internalise extracellular mitochondria and incorporate these organelles into the endogenous mitochondrial network.
Our findings suggest that several proteins are involved in fusion of exogenous mitochondria with endogenous mitochondria in iPS-CMs and HCFs. Mitofusin proteins are involved in tethering and fusing the mitochondrial outer membranes and OPA1 functions, in part, to enable inner mitochondrial membrane fusion34,35,36. Our results show that MFN1, MFN2, and OPA1 are expressed in both cardiomyocytes and fibroblasts and that a sizeable proportion of these proteins are associated with their respective mitochondria. Though mitofusin proteins concentrate at sites where exogenous and endogenous mitochondria are undergoing fusion, the presence of MFN1 and MFN2 beyond the boundaries of the mitochondrial network indicate these proteins may be involved in performing other cellular roles. In contrast, our immunoblot results reveal that OPA1 remains predominantly confined to the mitochondrial fraction and at least five OPA1 isoforms exist in cardiomyocytes. The long forms of OPA1 are membrane-anchored proteins involved in fusion of the mitochondrial inner membrane, whereas the short forms are soluble proteins associated with mitochondrial fission and other functions37.
To study fission-associated proteins in these cells, we examined the expression and post-translational modification of DRP1 by immunoblot analysis. DRP1 is a GTPase that oligomerises into structures that constrict and cleave mitochondria at specific sites on the outer membrane35. Serine phosphorylation of this protein regulates its activity38. For example, phosphorylation of DRP1 at S616 promotes mitochondrial fission during mitosis, whereas phosphorylation of S637 inhibits GTPase activity and fission39,40. Dephosphorylation of the latter amino acid promotes mitochondrial fission. The presence of fusion and fission proteins in iPS-CMs and HCFs is compatible with normal mitochondrial dynamics and our results confirm these cardiac cells contain the machinery necessary for fusion of exogenous mitochondria with the endogenous mitochondrial network. Given the metabolic and mitotic differences in these cells, it is not surprising to see variable expression and modification of these proteins. Interestingly, only HCFs expressed detectable levels of the mitophagy-related protein Parkin. Parkin is an E3 ubiquitin ligase that functions in a signaling pathway that identifies and eliminates dysfunctional mitochondria41,42. The absence of this protein in isolated mitochondrial lysates suggests there was little mitophagy occurring in either iPS-CMs or HCFs.
A complete understanding of the mechanisms that bring about endocytosis of extracellular mitochondria and fusion of those organelles with the endogenous mitochondrial network may provide a foundation for innovative new treatments to a number of human diseases and disorders43. Our earlier work has supported the use of mitochondrial transplantation to decrease infarction, increase ATP content, and improve functional performance of the ischemic heart6,7,8,9,17. We also demonstrated that delivery of exogenous mitochondria to the heart in animal models does not provoke an immune response nor produce arrhythmias7,21. Importantly, the clinical relevance and safety of autologous mitochondrial transplantation to the ischemic heart has recently been validated in five pediatric patients44. Aside from providing benefits to ischemic tissues, transplantation of exogenous mitochondria may prove to be useful for treating mitochondrial myopathies and a number of other diseases with underlying mitochondrial dysfunction13,15,16,17,18,20,45,46,47,48. Further study of the molecular mechanisms of exogenous mitochondrial endocytosis and fusion with endogenous mitochondria are warranted and these inquiries may guide development of innovative treatments directed at augmenting impaired mitochondria in a wide range of tissues and cells.