
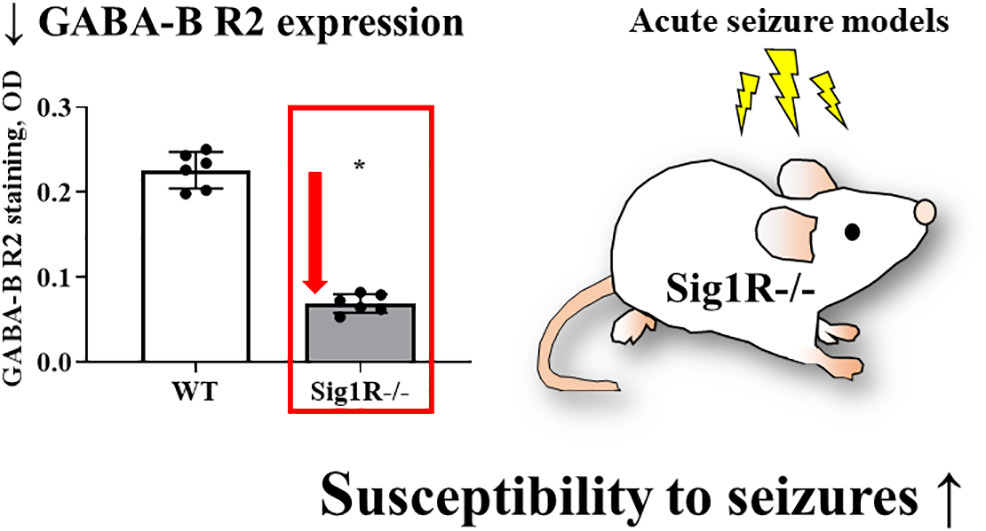
Genetic inactivation of the sigma-1 chaperone protein results in decreased expression of the R2 subunit of the GABA-B receptor and increased susceptibility to seizures
By Edijs Vavers, Baiba Zvejniece, Gundega Stelfa, Baiba Svalbe, Karlis Vilks, Einars Kupats, Rudolfs Mezapuke, Lasma Lauberte, Maija Dambrova, and Liga Zvejniece
Excerpt from the article published in Neurobiology of Disease, Volume 150, 2021, 105244, ISSN 0969-9961, https://doi.org/10.1016/j.nbd.2020.105244.
Editor’s Highlights
- The sigma-1 chaperone protein, also known as the sigma-1 receptor (Sig1R), in the cell plays a significant role in the modulation of the activity of endoplasmic reticulum-residing proteins
- Sig1R plays a pivotal role in the control of seizures.
- Sig1R antagonist-induced seizures resulted in Sig1R-independent downregulation of iNOS and GABA-Aγ2 receptor subunit genes.
- Sig1R behaves as an atypical auxiliary subunit to modulate potassium channel function.
- Sig1R and GABA-B receptor cooperation in the modulation of the seizure threshold.
- Sig1R is a significant molecular target for seizure modulation for the development of novel anti-seizure drugs.
Authors’ Highlights
- Sig1R−/− mice demonstrate decreased expression of the R2 subunit of the GABA-B receptor in the ventral part of the medial habenula.
- Sig1R−/− mice are more susceptible to pentylenetetrazol- and (+)bicuculline-induced seizures.
- Sig1R antagonist NE-100 demonstrates Sig1R-independent effects.
- NE-100-induced seizure severity is significantly less pronounced in Sig1R−/− mice.
- Sig1R is a valuable target for the development of novel anti-seizure drugs.
Abstract
There is a growing body of evidence demonstrating the significant involvement of the sigma-1 chaperone protein in the modulation of seizures. Several sigma-1 receptor (Sig1R) ligands have been demonstrated to regulate the seizure threshold in acute and chronic seizure models. However, the mechanism by which Sig1R modulates the excitatory and inhibitory pathways in the brain has not been elucidated. The aim of this study was to compare the susceptibility to seizures of wild type (WT) and Sig1R knockout (Sig1R−/−) mice in intravenous pentylenetetrazol (PTZ) and (+)-bicuculline (BIC) infusion-induced acute seizure and Sig1R antagonist NE-100-induced seizure models. To determine possible molecular mechanisms, we used quantitative PCR, Western blotting and immunohistochemistry to assess the possible involvement of several seizure-related genes and proteins. Peripheral tissue contractile response of WT and Sig1R−/− mice was studied in an isolated vasa deferentia model. The most important finding was the significantly decreased expression of the R2 subunit of the GABA-B receptor in the hippocampus and habenula of Sig1R−/− mice. Our results demonstrated that Sig1R−/− mice have decreased thresholds for PTZ- and BIC-induced tonic seizures. In the NE-100-induced seizure model, Sig1R−/− animals demonstrated lower seizure scores, shorter durations and increased latency times of seizures compared to WT mice. Sig1R-independent activities of NE-100 included downregulation of the gene expression of iNOS and GABA-A γ2 and inhibition of KCl-induced depolarization in both WT and Sig1R−/− animals. In conclusion, the results of this study indicate that the lack of Sig1R resulted in decreased expression of the R2 subunit of the GABA-B receptor and increased susceptibility to seizures. Our results confirm that Sig1R is a significant molecular target for seizure modulation and warrants further investigation for the development of novel anti-seizure drugs.
Graphical abstract
1. Introduction
Epilepsy is a chronic neurological disease that is characterized by seizures. The seizures could be caused by disturbances at the receptor function and neurotransmission levels, metabolic problems in the central nervous system or some conditions leading to pathological changes in the brain, such as traumatic brain injuries, stroke, status epilepticus or infectious diseases (Alyu and Dikmen, 2017). For the treatment of seizures, heterogeneous groups of antiepileptic drugs(AEDs) are used. However, a number of patients do not fully respond to currently available treatment, and existing AEDs achieve seizure freedom only in approximately two-thirds of patients (Perucca et al., 2018). Therefore, searching for new possible mechanisms and novel AEDs is highly necessary to identify effective treatments for patients with uncontrolled seizures.
The sigma-1 chaperone protein, also known as the sigma-1 receptor (Sig1R), in the cell plays a significant role in the modulation of the activity of endoplasmic reticulum-residing proteins, as well as nuclear, mitochondrial and plasma-membrane proteins, which are in close proximity to the endoplasmic reticulum(Mavlyutov et al., 2015; Su et al., 2016). It has been shown that sigma ligands are involved in modulation of the seizure threshold, and both anti-convulsive and pro-convulsive activity has been observed for both agonists and antagonists (Vavers et al., 2017). Our previous data showed that the Sig1R antagonist NE-100 demonstrated pro-convulsive activity at a dose of 25 mg/kg after i.p. administration in a PTZ-induced seizure model, and after administration at doses of 50 and 75 mg/kg, NE-100 induced convulsions in naïve animals (Vavers et al., 2017). On the other hand, positive allosteric modulators of Sig1R clearly demonstrated anti-convulsive activity in in vivo seizure models (Guo et al., 2015; reviewed in Vavers et al., 2019). Taken together, these findings strongly suggest that Sig1R could be used as a novel drug target to treat seizures. The mechanism governing how Sig1R can control the balance between excitation and inhibition in vivo has not been fully elucidated. The first Sig1R knockout (Sig1R−/−) mice were described in 2003 (Langa et al., 2003), and to date, no spontaneous seizures have been observed in Sig1R−/− mice.
In several in vitro studies, it has been shown that Sig1R could alter neuronal excitability by modulating the function of voltage-gated sodium, potassium and calcium channels (Kourrich, 2017). The activation of Sig1R has been demonstrated to inhibit glutamate release in rat cerebral cortex nerve endings (Lu et al., 2012). On the other hand, it has been shown that endogenous activators and agonists of Sig1R, such as pregnenolone sulfate, dehydroepiandrosterone sulfate and (+)-pentazocine, can increase presynaptic glutamate release in rat hippocampal neurons (Meyer et al., 2002; Dong et al., 2007). In addition, pregnenolone sulfate and dehydroepiandrosterone sulfate have been demonstrated to negatively modulate and decrease the activity of the GABA-A receptor (Maurice et al., 2001) and thus may increase the sensitivity of mice to pentylenetetrazol (PTZ)-induced seizures (Reddy and Kulkarni, 1998). The lack of Sig1R in Sig1R−/− mice has been associated with dysfunction of GABA-A receptor-mediated inhibition in basolateral amygdalaprincipal neurons, probably through the reduction of NMDA receptor-mediated GABA release (Zhang et al., 2017). A bidirectional modulation of Sig1R agonists of NMDA receptor evoked excitatory transmission, in which a low dose of Sig1R agonist is excitatory and a high dose is inhibitory (Liang and Wang, 1998), suggesting a complex mechanism that is responsible for regulating the balance between excitation and inhibition by Sig1R.
To determine the possible mechanisms governing the role of Sig1R in the control of seizures, the aim of this study was to compare the seizure threshold and behavioral response of WT and Sig1R−/− mice in different acute seizure models in vivo. Changes in seizure-related gene and protein expression were analyzed using PCR, Western blotting and immunohistochemistry techniques. The isolated vasa deferentia model was used to test the role of Sig1R in electrical current-induced contractility and potassium chloride-induced depolarization.
3. Results
3.1. Sig1R−/− mice are more prone to PTZ- and BIC-induced tonic seizures
The susceptibility to seizures of WT and Sig1R−/− mice was compared in timed intravenous PTZ and BIC infusion-induced acute seizure models. The 1% PTZ intravenous infusion induced clonic and tonic seizures in WT animals at doses of 33 ± 1 mg/kg and 70 ± 6 mg/kg, respectively (Fig. 1A, B). Compared to WT mice, Sig1R−/− mice demonstrated clonic seizures at a dose of 31 ± 1 mg/kg of PTZ (Fig. 1A), while tonic seizures were observed at a dose of 50 ± 4 mg/kg of PTZ (Fig. 1B). The response of the PTZ-induced clonic seizures did not differ between WT and Sig1R−/− animals (Mann-Whitney U test: p = 0.2328; Fig. 1A). In Sig1R−/− mice, the PTZ-induced tonic seizure threshold was significantly decreased by 28% compared to WT animals (Mann-Whitney U test: p = 0.0041; Fig. 1B). BIC-induced clonic and tonic seizures in WT animals were observed at doses of 0.61 ± 0.03 mg/kg and 1.12 ± 0.09 mg/kg of BIC, respectively (Fig. 2A and B). The response of the BIC-induced clonic seizures did not differ between WT and Sig1R−/− animals (unpaired t-test: t(17) = 0.8, p = 0.4539; Fig. 2A). Similar to the PTZ-induced seizure model, the tonic seizure threshold was significantly decreased in Sig1R−/− mice by 22% (0.88 ± 0.04 mg/kg of BIC; unpaired t-test: t(17) = 2.6, p = 0.0187) compared to WT mice (Fig. 2B), thereby demonstrating that Sig1R−/− mice are more susceptible to tonic seizures in both PTZ- and BIC-induced seizure models.
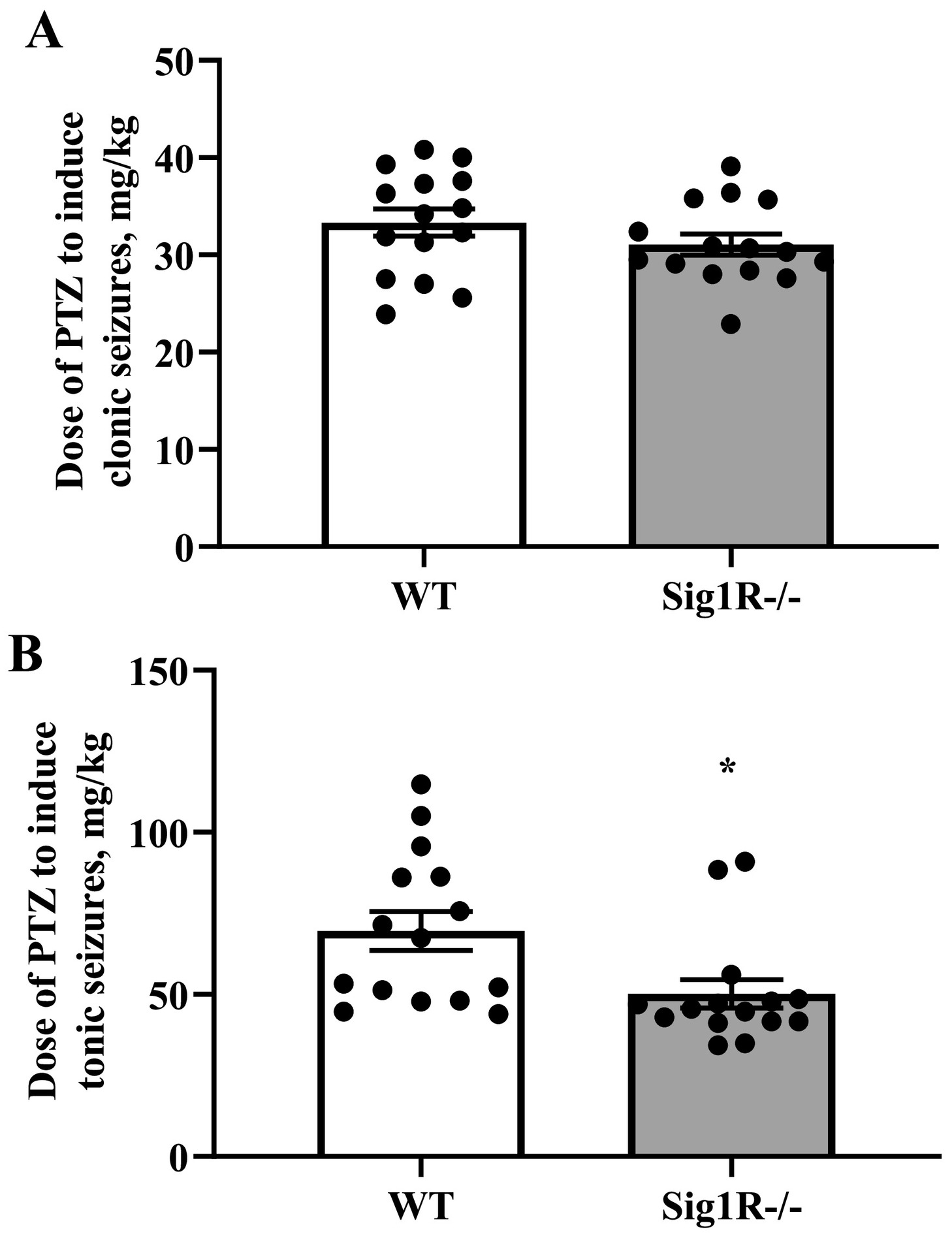
Comparison of the response of WT and Sig1R−/− mice in the PTZ-induced seizure model. PTZ-induced clonic (A) and tonic (B) seizure thresholds. Data are expressed as the mean ± SEM (n = 15). *p < 0.05 Sig1R−/− mice vs WT mice (Mann-Whitney test).
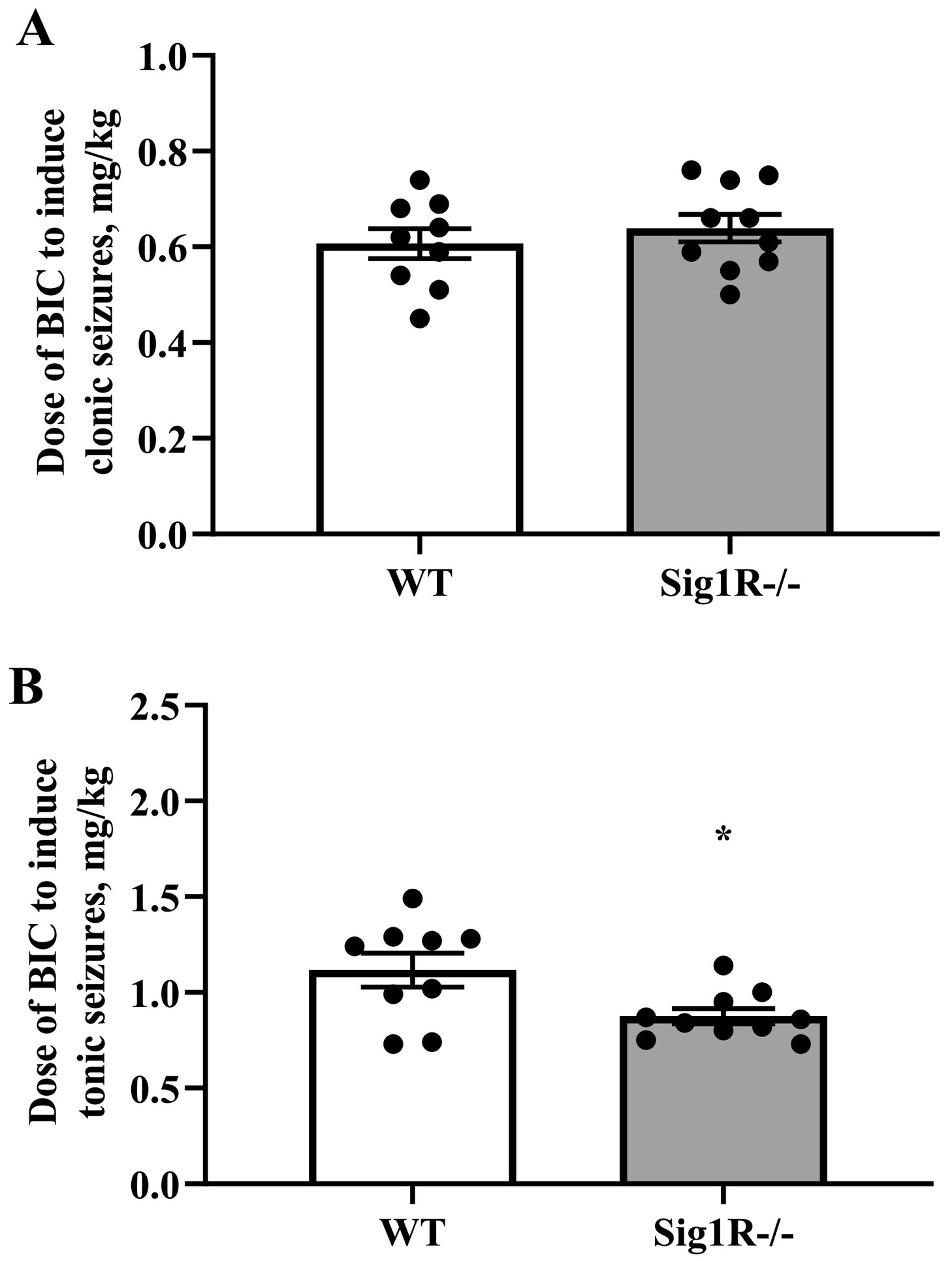
Comparison of the response of WT and Sig1R−/− mice in the BIC-induced seizure model. BIC-induced clonic (A) and tonic (B) seizure thresholds. Data are expressed as the mean ± SEM (n = 9 WT mice and n = 10 Sig1R−/− mice). *p < 0.05 Sig1R−/− mice vs WT mice (unpaired t-test).
3.2. Sig1R−/− mice demonstrate decreased severity of seizure in the NE-100-induced seizure model
The responses of WT and Sig1R−/− mice in the NE-100-induced seizure model were different and could be divided into two phases. Acute seizures of NE-100 (up to 20 min) were observed in both WT and Sig1R−/− mice, while the second phase, which started 20–22 min after administration of NE-100, was observed only in WT animals (Fig. 3A). The average maximal seizure score in WT mice was 7.6 while in Sig1R−/− mice it was 5.4 points (Fig. 3A). There was a significant genotype x time interaction observed between seizure severities (two-way repeated-measures ANOVA: F(59, 944) = 4.3, p < 0.0001 for genotype x time interaction; F(1, 16) = 17.4, p = 0.0007 for genotype; F(5, 83) = 19.2, p < 0.0001 for time; Fig. 3A). As demonstrated by the data expressed as the areas under the curves (AUCs), the seizure severity after administration of NE-100 in Sig1R−/− mice was less pronounced than that in WT animals (Mann-Whitney U test: p = 0.0040; Fig. 3B). Compared to WT animals, in which seizures lasted at least 34 min (Fig. 3A, C), the NE-100-induced seizures in Sig1R−/− mice lasted for significantly shorter lengths of time (Mann-Whitney U test: p = 0.0012; Fig. 3C). One Sig1R−/− mouse did not show any signs of seizures after administration of NE-100 at a dose of 75 mg/kg (Fig. 3B, C). Sig1R−/− mice exhibited a significantly higher latency time to NE-100-induced seizures (Mann-Whitney Utest: p = 0.0380; Fig. 3D). NE-100-induced clonic seizures in Sig1R−/− mice were observed for a significantly shorter time than in WT animals (Mann-Whitney U test: p = 0.0199 for clonic seizures while lying on the belly; p = 0.0097 for clonic seizures while lying on the side; Fig. 4).
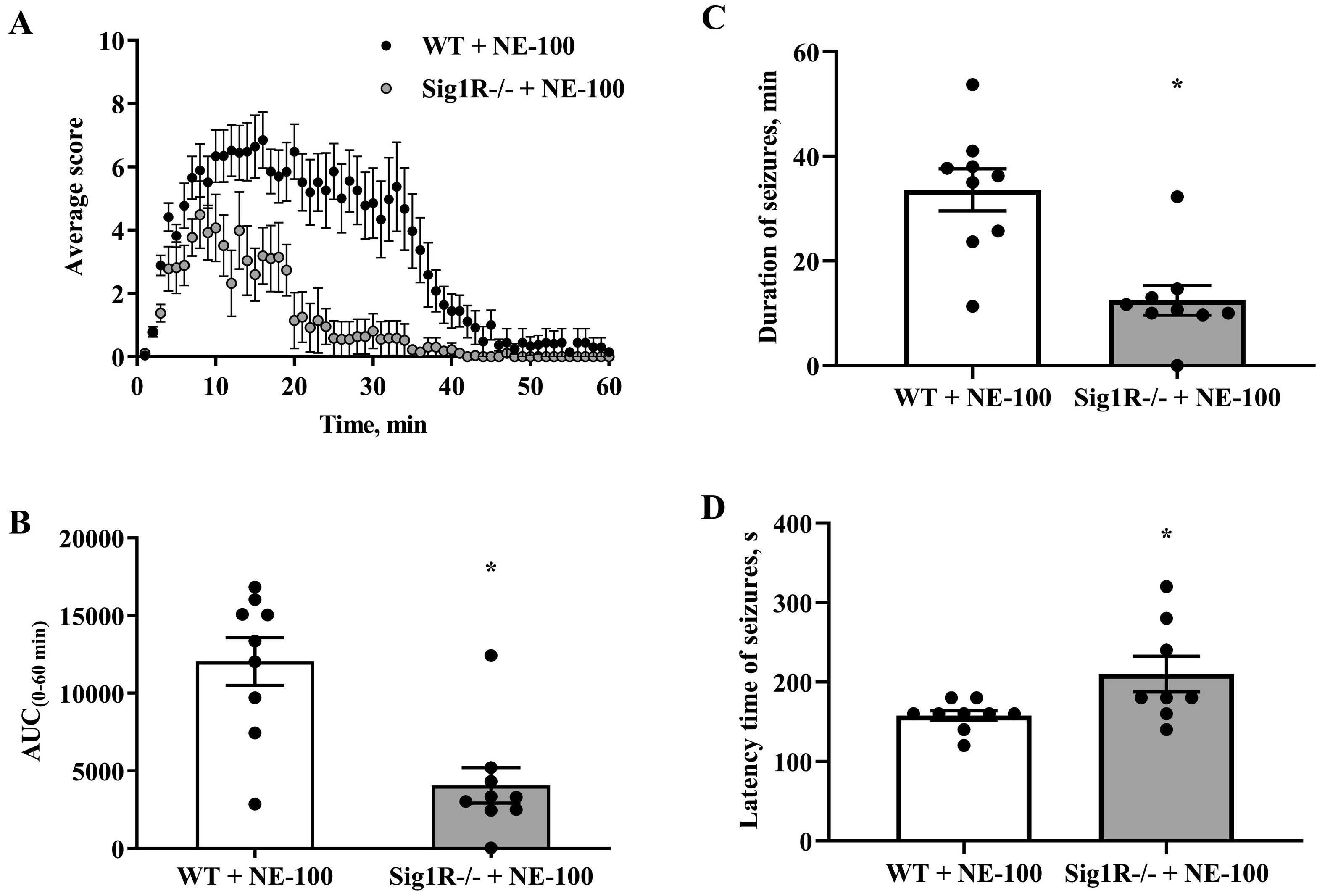
NE-100-induced acute seizure model. (A) Average behavioral scores for each group during the 60-min observation period. Data are expressed as the mean for each 1 min period. (B) The area under the curve (AUC0-60min) was calculated from the behavioral scoring curve. (C) Duration time of seizures for each animal and (D) latency time of NE-100-induced seizures. Data are expressed as the mean ± SEM (n = 9). *p < 0.05 Sig1R−/− mice vs WT mice (Mann-Whitney test).
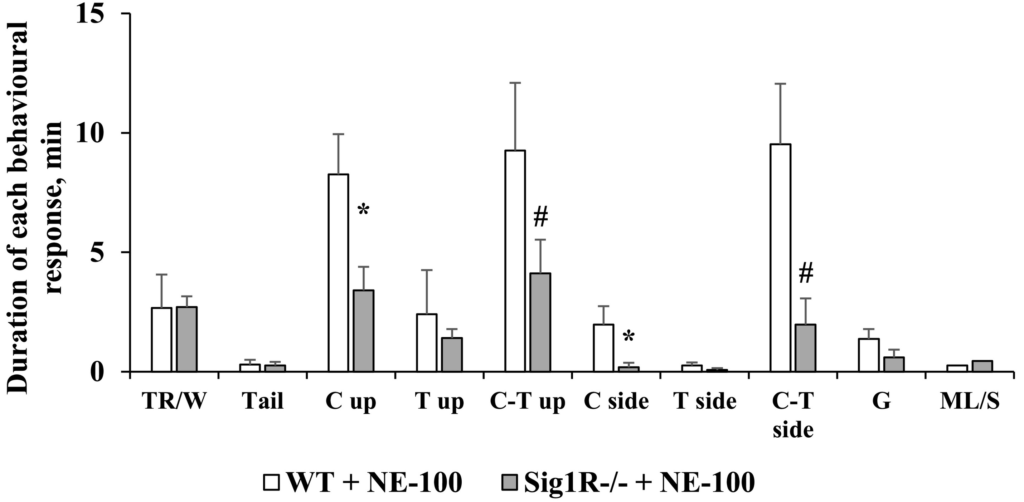
Comparison of the duration of each observed seizure behavior after administration of NE-100 in WT and Sig1R−/− mice. The different behavioral categories include: trembling or wobbly gait (TR/W), tail lifting (Tail), clonic, tonic and clonic-tonic seizures while lying on the belly (C, T and C-T up, respectively), clonic, tonic and clonic-tonic seizures while lying on the side (C, T and C-T side, respectively), generalized seizures with wild jumping and running (G), and motionless or sleeping (ML/S). Data are expressed as the mean ± SEM (n = 9). *p < 0.05 and #p = 0.06 Sig1R−/− mice vs WT mice (Mann-Whitney test).
3.3. NE-100-induced seizures result in the downregulation of mRNA levels of iNOS and the γ2 subunit of the GABA-A receptor
Sig1R gene expression was observed only in WT mice, but not in Sig1R−/− mice (Table 1). No significant differences in other investigated gene expression levels were found between WT and Sig1R−/− control mice (Table 1). However, Sig1R−/− animals presented a slight tendency of decreased GABA-B R2 mRNA levels (Mann-Whitney U test: p = 0.1111; Table 1). The administration of NE-100 induced significant downregulation of the gene expression of iNOS (Mann-Whitney U test: p = 0.0022) and the γ2 subunit of the GABA-A receptor (Mann-Whitney U test: p = 0.0303) in WT mice (Table 1). Significant downregulation of the gene expression of iNOS (Mann-Whitney U test: p = 0.0159) and the γ2 subunit of the GABA-A receptor (Mann-Whitney U test: p = 0.0286) was also observed in Sig1R−/− mice after administration of NE-100 (Table 1).
Table 1. Gene expression.
| Genes | Protein | Gene expression, fold change vs WT | ||
|---|---|---|---|---|
| Sigmar1 | Sig1R | – | 0.8 ± 0.2 | – |
| Pgrmc1 | Sig2R | 1.1 ± 0.6 | 1.0 ± 0.2 | 0.9 ± 0.1 |
| Grin1 | NMDAR1 | 1.0 ± 0. | 0.9 ± 0.3 | 0.8 ± 0.2 |
| Drd1 | D1R | 0.8 ± 0.4 | 1.1 ± 0.8 | 1.0 ± 0.5 |
| Drd2 | D2R | 0.8 ± 0.5 | 1.0 ± 0.8 | 1.0 ± 0.6 |
| Nos2 | iNOS | 1.0 ± 0.5 | 0.5 ± 0.2 ↓* | 0.5 ± 0.1 ↓* |
| Gabra5 | GABA-A α5 | 1.0 ± 0.8 | 1.1 ± 0.3 | 0.8 ± 0.2 |
| Gabrb3 | GABA-A β3 | 1.1 ± 0.6 | 0.9 ± 0.2 | 0.8 ± 0.1 |
| Gabrg2 | GABA-A γ2 | 0.9 ± 0.7 | 0.4 ± 0.4 ↓* | 0.5 ± 0.3 ↓* |
| Gabbr1 | GABA-B R1 | 0.9 ± 0.1 | 0.9 ± 0.3 | 0.9 ± 0.2 |
| Gabbr2 | GABA-B R2 | 0.6 ± 0.3 ↓ | 1.3 ± 0.6 | 1.2 ± 0.3 |
| HSPA5 | BiP | 0.9 ± 0.2 | 1.1 ± 0.4 | 0.8 ± 0.4 |
| P2rx1 | P2RX1 | 0.7 ± 0.5 | 0.7 ± 0.4 | 1.1 ± 0.3 |
| Ryr3 | RyR3 | 0.9 ± 0.2 | 1.1 ± 0.6 | 0.7 ± 0.2 |
| Kcnj3 | Kir3.1 | 0.8 ± 0.1 | 0.9 ± 0.4 | 1.0 ± 0.6 |
| Kcnj9 | Kir3.3 | 0.7 ± 0.1 | 0.6 ± 0.3 ↓ | 0.8 ± 0.3 |
| Aif1 | Iba1 | 1.1 ± 0.4 | 1.3 ± 0.8 | 1.0 ± 0.3 |
| Cd68 | CD68 | 1.2 ± 0.6 | 1.1 ± 0.1 | 0.8 ± 0.2 |
3.4. Sig1R−/− mice demonstrate reduced expression of the R2 subunit of the GABA-B receptor
Based on gene expression data, we attempted to evaluate the protein expression levels of the R2 subunit of the GABA-B receptor and the γ2 subunit of the GABA-A receptor in the brain between WT and Sig1R−/− mice by using Western blot analysis. Sig1R−/− mice demonstrated significantly decreased expression of the GABA-B receptor R2 subtype compared to WT mice (Fig. 5A and B). The results showed no significant differences in the expression of the γ2 subunit of the GABA-A receptor between WT and Sig1R−/− animals (Fig. 5A and C). Although NE-100 induced a significant downregulation of the expression of the γ2 subunit of the GABA-A receptor, no difference in protein levels was observed in WT mice (Supplementary Fig. 1A and Supplementary Fig. 1B) and Sig1R−/− mice (Supplementary Fig. 1C and Supplementary Fig. 1D) after administration of NE-100. In addition, NE-100 was not observed to bind directly to the GABA-A receptor in the [3H]muscimol binding assay (Supplementary Fig. 2).
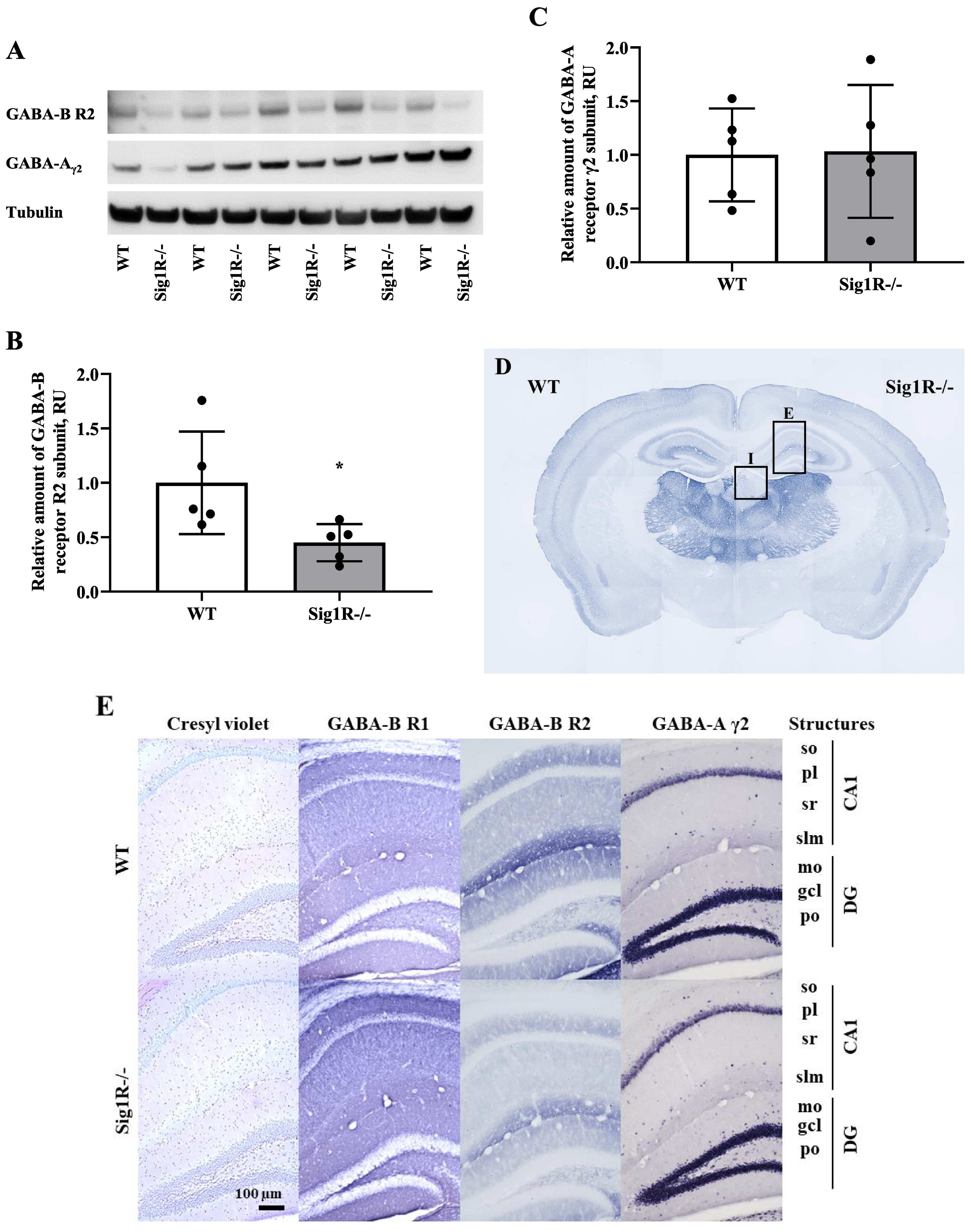
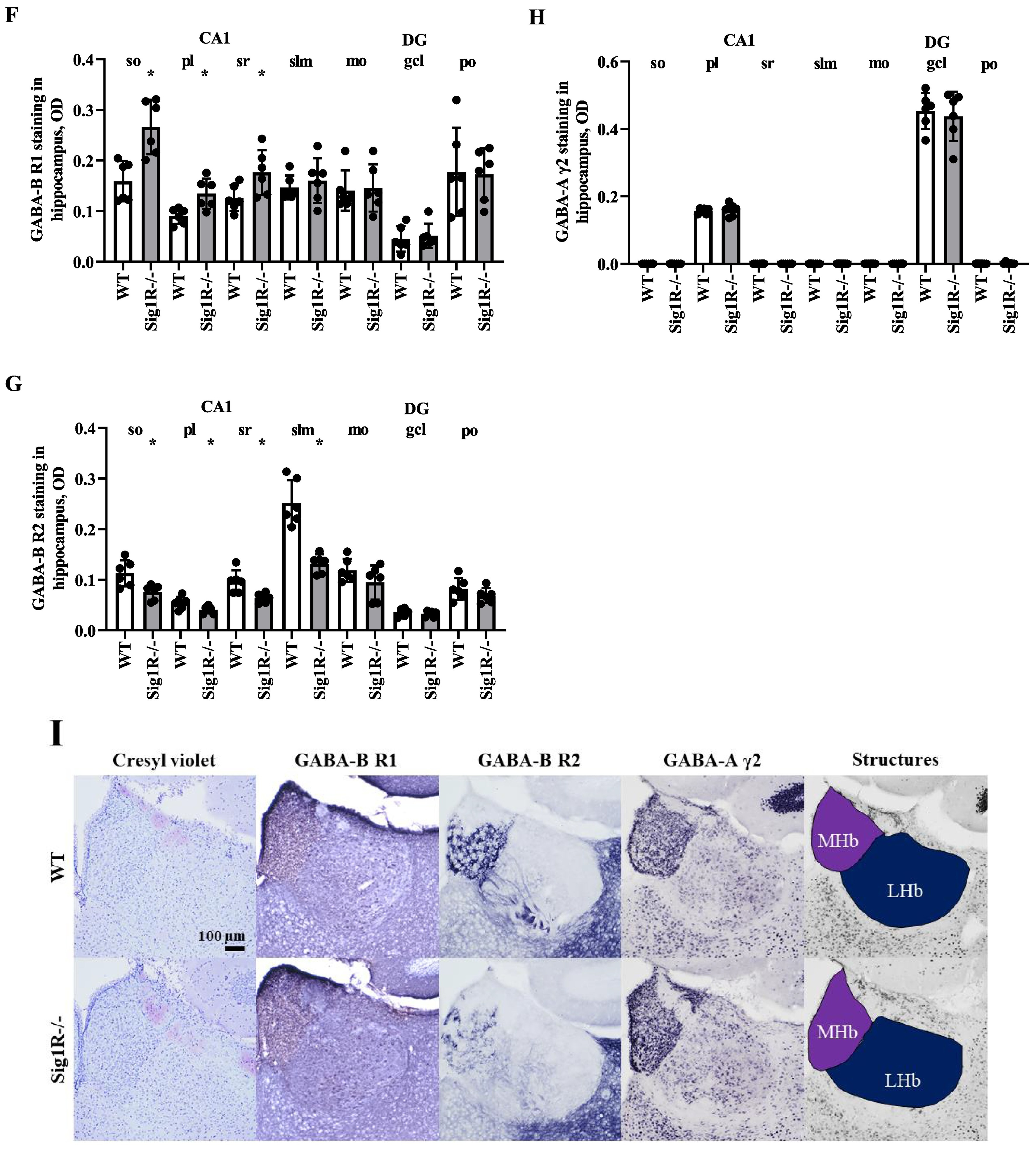
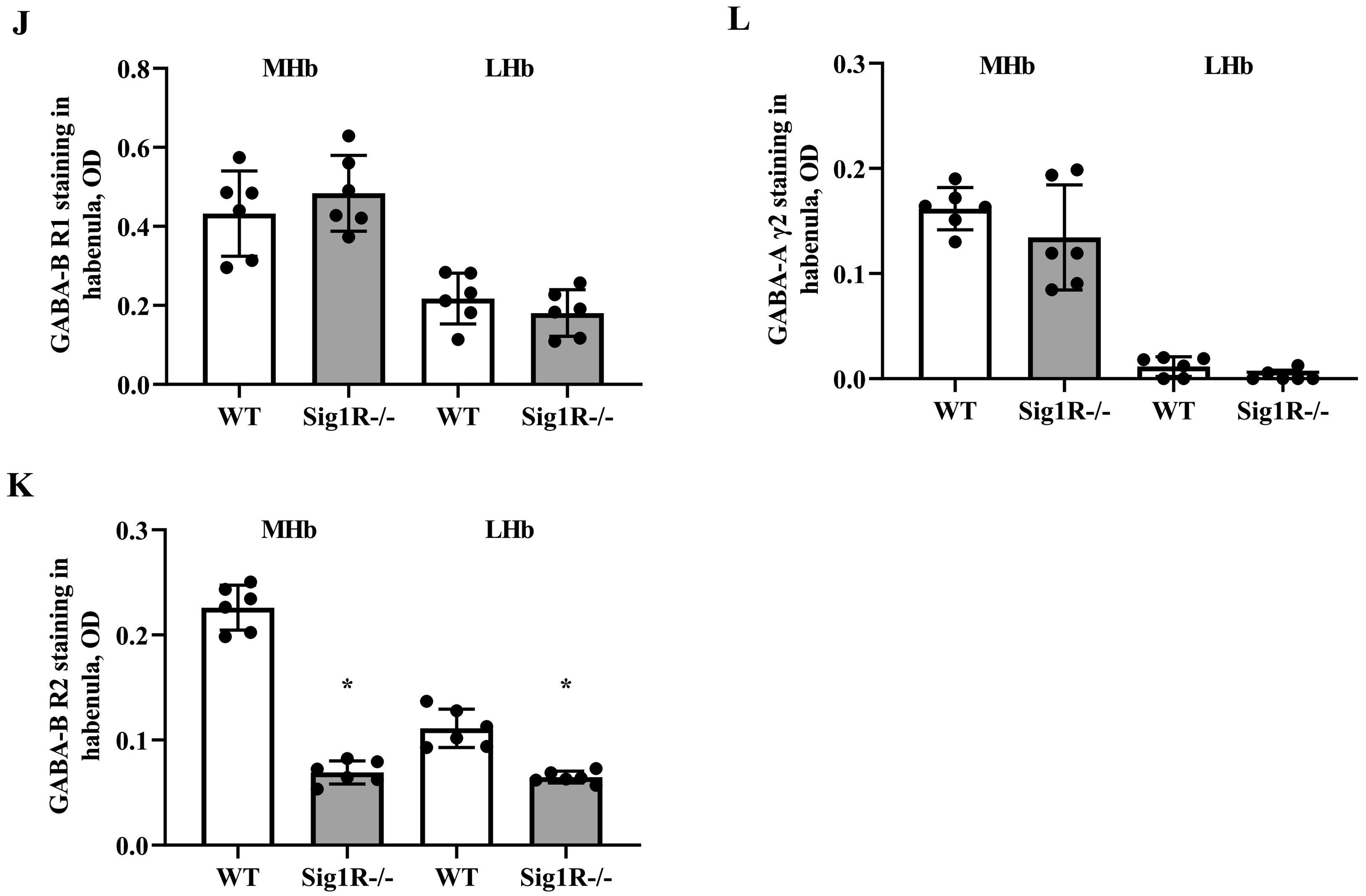
GABA-A and GABA-B receptor expression in WT and Sig1R−/− mouse brains. Western blot images (A) and calculated relative amounts of the R2 subunit of the GABA-B (B) and γ2 subunit of the GABA-A (C) receptors of WT vs Sig1R−/− control animals. Data are expressed as the mean ± SD (n = 5). *p < 0.05 Sig1R−/− mice vs WT mice (Mann-Whitney U test). Overview of GABA-B R2 staining in WT (on the left) and Sig1R−/− (on the right) mouse brain hemispheres (D) with the indicated areas used for analysis. Representative images of immunohistochemical staining in the hippocampus (E) and habenula (I). Measured optical density (OD) of the staining intensity of the GABA-B R1 in the hippocampus (F), and habenula (J), GABA-B R2 in the hippocampus (G) and habenula (K) and GABA-A γ2 in the hippocampus (H) and habenula (L). Data are expressed as the mean ± SD (n = 6). *p < 0.05 Sig1R−/− mice vs WT mice (Mann-Whitney U test). Cornu Ammonis 1 (CA1), dentate gyrus (DG), stratum oriens (so), pyramidal layer(pl), stratum radiatum (sr), stratum lacunosum-moleculare (slm), molecular layer (mo), granule celllayer (gcl), polymorph layer (po), medial habenula (MHb) and lateral habenula (LHb).
To test which brain regions have decreased expression of the GABA-B receptor in Sig1R−/− mice, immunohistochemical staining of both the R1 and R2 subunits of the GABA-B receptor was performed (Fig. 5D, E and I). Sig1R−/− mice demonstrated significantly increased staining of GABA-B R1 in the stratum oriens (Mann-Whitney U test: p = 0.0043) and stratum radiatum (Mann-Whitney U test: p = 0.0260) while reduced staining of GABA-B R2 was observed in the stratum oriens (Mann-Whitney U test: p = 0.0108), stratum radiatum (Mann-Whitney U test: p = 0.0087) and stratum lacunosum-moleculare (Mann-Whitney U test: p = 0.0022) of the hippocampal Cornu Ammonis 1 (CA1) region (Fig. 5G). The most marked difference in the staining of the R2 subunit of the GABA-B receptor in Sig1R −/− mice was found in the habenula(Fig. 5I). The expression of the R2 subunit of the GABA-B receptor was significantly decreased in the Sig1R −/− mice medial and lateral habenula (Mann-Whitney U test: p = 0.0022 and p = 0.0476, respectively; Fig. 5K). Immunofluorescence staining of the GABA-B R2 in the medial habenula confirmed DAB staining results (Mann-Whitney U test: p = 0.0022; Fig. 6A and B). By using co-labeling experiments the most significant decrease of the R2 staining of the GABA-B receptor in Sig1R−/− mice was found in the ventral part of the medial habenula (Fig. 6C and D) and was confirmed with confocal imaging (Fig. 6E). GABA-B R1 staining in the habenula did not differ between WT and Sig1R−/− animals (Fig. 5J and Fig. 6E). Administration of NE-100 did not change the GABA-B R2 protein levels in the hippocampus (Supplementary Table 2) and habenula (Supplementary Fig. 4A and 4B) of WT and Sig1R−/− mice. We did not observe any significant difference in the staining of the γ2 subunit of the GABA-A receptor in the brain between WT and Sig1R−/− mice (Fig. 5E, H, I and L).
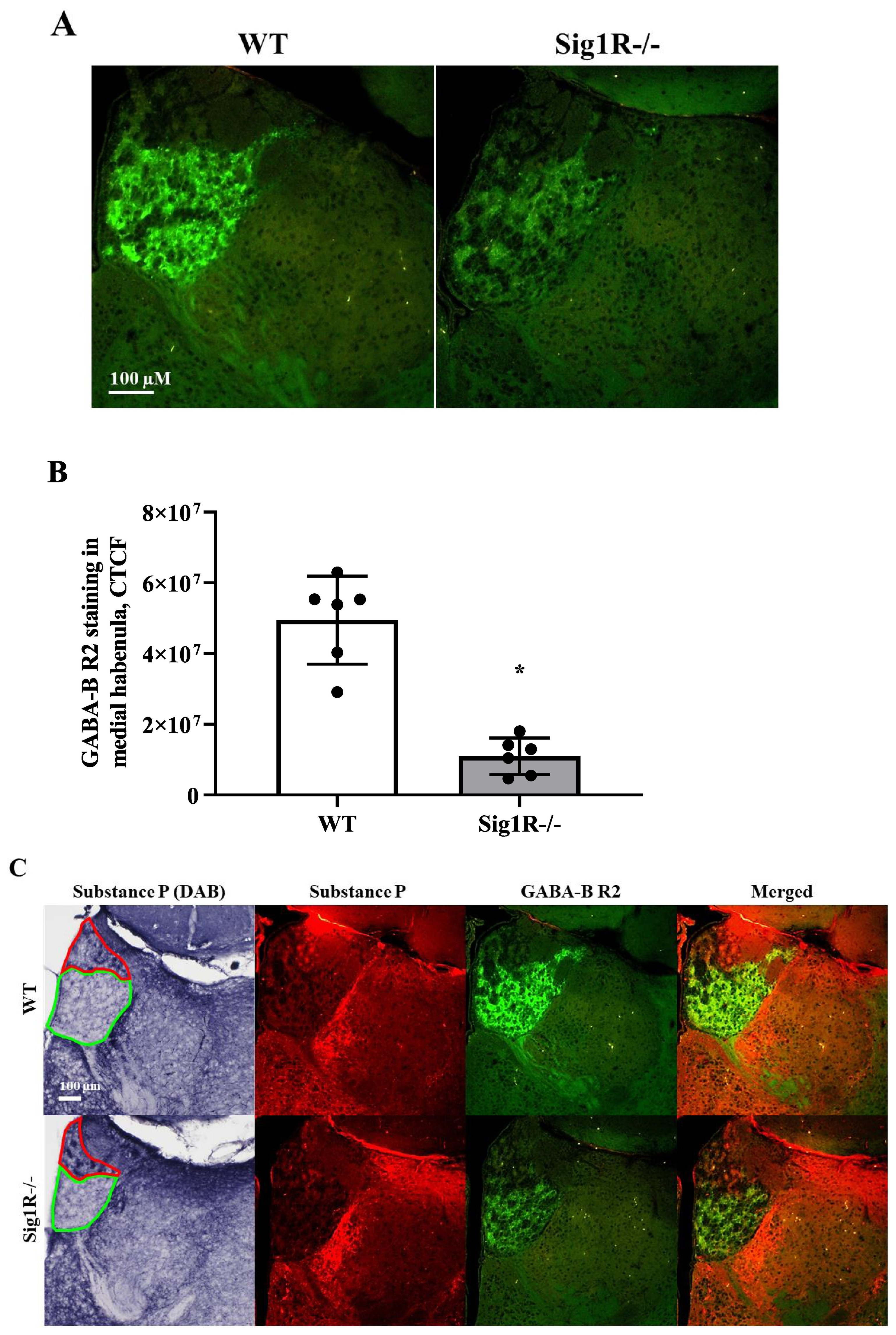
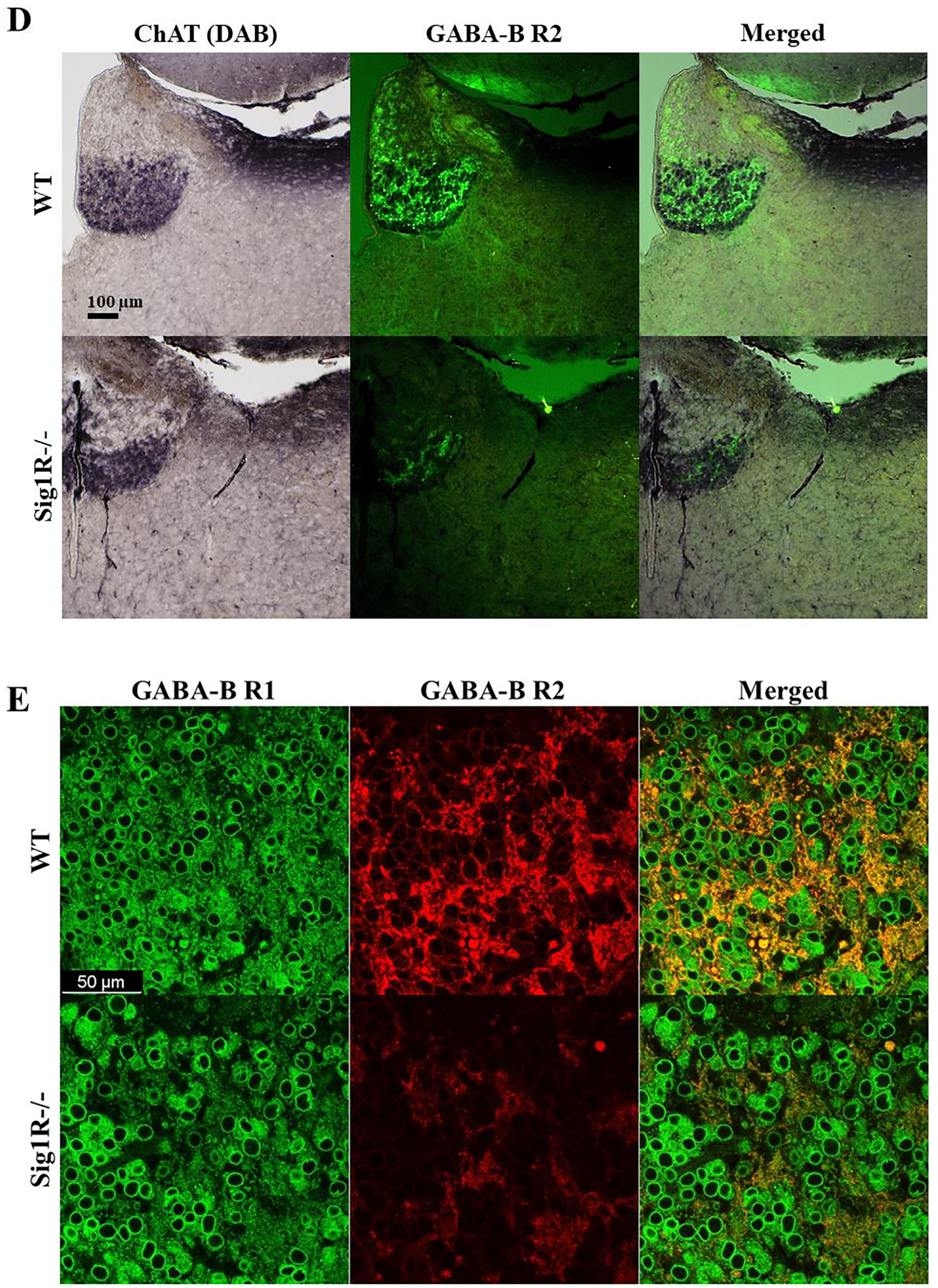
Immunofluorescence of the R2 subunit of GABA-B receptors in the medial habenula. (A) Comparison of GABA-B R2 expression in WT and Sig1R−/− mice by using immunofluorescence, (B) measured corrected total cell fluorescence (CTCF). Data are expressed as the mean ± SD (n = 6). *p < 0.05 Sig1R−/− mice vs WT mice (Mann-Whitney U test). (C) DAB and immunofluorescence staining of Substance P (a marker of the dorsal part of the medial habenula, red) and double immunostaining with R2 subunit of GABA-B receptors (green). Red line demonstrates dorsal part and green line demonstrates ventral part of the medial habenula. (D) Combined DAB and immunofluorescence staining of ChAT (a marker of the ventral part of the medial habenula) and GABA-B R2 (green), respectively. (E) A representative confocal image demonstrating colocalization of R1 subunit (green) and R2 subunit (red) of the GABA-B receptor in the ventral part of the medial habenula of WT and Sig1R−/− mice.
3.5. NE-100 enhances electrically stimulated contractions of isolated vasa deferentia and inhibits KCl-induced maximal contractility
The isolated vasa deferentia model has been used previously to test and describe the activity of Sig1R ligands. The activity of NE-100 on electrical field stimulation-induced contractions of isolated vasa deferentia of WT and Sig1R−/− mice was investigated. NE-100 dose-dependently enhanced electrical current-induced contractions of the vasa deferentia of WT and Sig1R −/− mice (Fig. 7A). A two-way repeated measures ANOVA showed a significant interaction between concentration of NE-100 and genotype (F(3,33) = 43.9, p < 0.0001) and main effects of concentration of NE-100 (F(1, 16) = 98.6, p < 0.0001) and genotype (F(1,11) = 17.8, p = 0.0015; Fig. 7A). The contractions of vasa deferentia were significantly higher in WT than in Sig1R−/− mice after treatment with NE-100 at concentrations of 3 and 10 μM (two-way repeated measures ANOVA followed by Bonferroni test: p < 0.0001 and p < 0.0001, respectively; Fig. 7A). NE-100 significantly inhibited the response of vasa deferentia to KCl in both WT and Sig1R−/− animals (one-way ANOVA: F(3,22) = 56.7, p < 0.0001; Fig. 7B). Compared to WT mice, Sig1R−/− mice demonstrated decreased maximal contractility of vasa deferentia, which was measured by the addition of 100 mM KCl (one-way ANOVA followed by Tukey’s test: p = 0.0011; Fig. 7B). We examined the dose-dependent activity of barium chloride, a nonspecific inhibitor of inward-rectifier potassium channels, on the contractility of vasa deferentia isolated from WT and Sig1R−/− animals (Fig. 7C). Barium chloride dose-dependently enhanced electrical current-induced contractions of both genotype vasa deferentia (two-way repeated-measures ANOVA: F(5, 50) = 179.3, p < 0.0001 for concentration of barium chloride; Fig. 7C). The effect of barium chloride was significantly less pronounced on vasa deferentia isolated from Sig1R−/− mice (two-way repeated measures ANOVA: F(5, 50) = 10.1, p < 0.0001 for concentration of barium chloride and genotype interaction; Fig. 7C).
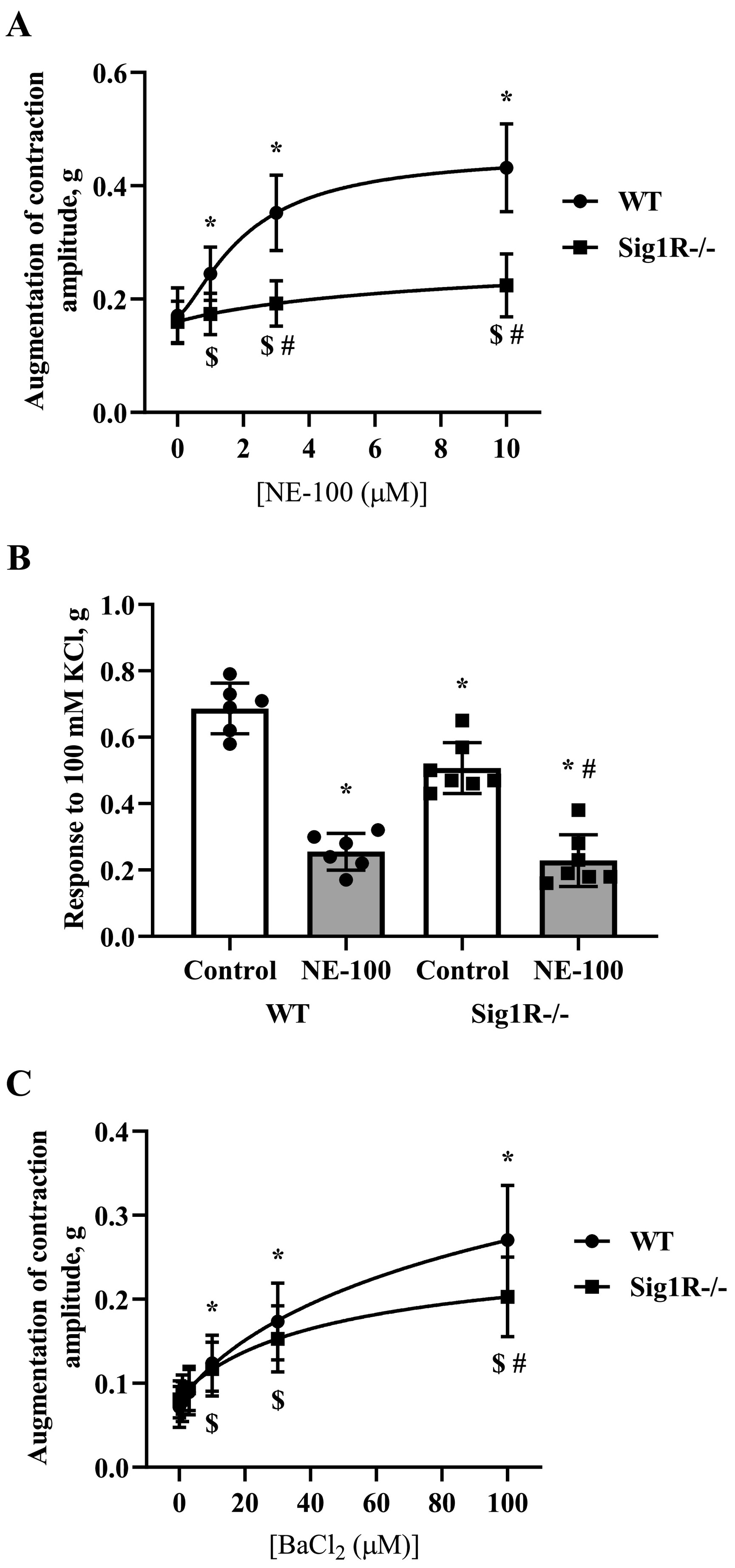
Isolated vasa deferentia model. (A) NE-100-enhanced electrical current-induced contractions of vasa deferentia. Data are shown as the mean ± SD (n = 6 WT, n = 7 Sig1R−/− mice). *p < 0.05 dose-dependent effect of NE-100 vs baseline of WT mice, $p < 0.05 dose-dependent effect of NE-100 vs baseline of Sig1R−/− mice (two-way repeated measures ANOVA followed by Dunnett’s test for comparison with baseline within each group); #p < 0.05 WT vs Sig1R−/− (two-way repeated measures ANOVA followed by Bonferroni test for comparison of NE-100 effect between genotypes). (B) The effect of 10 μM NE-100 on KCl-induced maximal contractility. Data are shown as the mean ± SD (n = 6 WT, n = 7 Sig1R−/− mice). *p < 0.05 vs WT control; #p < 0.05 vs Sig1R−/− control (one-way ANOVA followed by Tukey’s test). (C) The effect of barium chloride, a nonspecific inhibitor of inward-rectifier potassium channels, on the contractility of vasa deferentia. Data are shown as the mean ± SD (n = 6 WT, n = 6 Sig1R−/− mice). *p < 0.05 dose-dependent effect of barium chloride vs baseline of WT mice, $p < 0.05 dose-dependent effect of barium chloride vs baseline of Sig1R−/− mice (two-way repeated measures ANOVA followed by Dunnett’s test for comparison with baseline within each group); #p < 0.05 WT vs Sig1R−/− (two-way repeated measures ANOVA followed by Bonferroni test for comparison of barium chloride effect between genotypes).
4. Discussion
This study demonstrates a reduced tonic seizure threshold of Sig1R−/− animals in PTZ and BIC-induced acute seizure models and decreased expression of the R2 subunit of the GABA-B receptor in the ventral part of the medial habenula and CA1region of hippocampus compared to that of age-matched WT mice. NE-100-induced seizure severity was significantly less pronounced in Sig1R−/− mice. Sig1R-independent downregulation of gene expression of iNOS and GABA-Aγ2 in the brain was observed 75 min after the administration of NE-100. Sig1R−/− animals showed a reduced response to KCl, which resulted in decreased maximal contractility of vasa deferentia compared to WT animals. NE-100 inhibited KCl-induced depolarization of the vasa deferentia in both WT and Sig1R−/− mice.
Since Sig1R−/− mice previously exhibited dysfunction of GABA-A receptor-mediated inhibition (Zhang et al., 2017), to compare the seizure threshold in WT and Sig1R−/− animals, we used PTZ and BIC, which both inhibit the GABA-A receptor and induce seizures in naïve animals (Galanopoulou and Moshé, 2014). Our results showed that Sig1R−/− mice are more vulnerable to PTZ- and BIC-induced tonic seizures. To date, direct protein-protein interactions of Sig1R with the GABA receptors have not been demonstrated. Previously, the analysis of RT-PCR samples from C57Bl/6 background mice showed that the levels of GABA-Aα4 and GABA-Aδ mRNA in brain samples from the basolateral amygdala did not differ significantly between Sig1R−/− and WT mice (Zhang et al., 2017). In addition, we did not find differences in GABA-Aα5, GABA-Aβ3 and GABA-Aγ2 mRNA levels in CD-1 background Sig1R−/− mouse brain samples. Western blotting results showed that the γ2 subunit of GABA-A receptor expression in brain samples was not altered in Sig1R−/− mice compared to WT animals. It was reported previously that the direct function of the GABA-A receptor in hippocampal pyramidal neurons of Sig1R−/− mice is not changed (Sha et al., 2013). Therefore, different seizure thresholds in PTZ- and BIC-induced seizure models could not be associated with different amounts or functions of the GABA-A receptor in CD-1 background WT and Sig1R−/− mice.
In our study, we demonstrated significantly decreased expression of the R2 subunit of the GABA-B receptor in the habenula and hippocampus of Sig1R−/− mouse brain tissue. We found significantly increased expression of R1 subunit of GABA-B receptors in the CA1 region of the hippocampus of Sig1R−/− mice. This could be considered as a compensatory mechanism in Sig1R−/− animals due to decreased expression of the R2 subunit of the GABA-B receptor in the structures of CA1. Altered GABA-B receptor function has been associated with a variety of neurological and psychiatric disorders, including epilepsy, depression, anxiety, cognition, nociception and drug addiction (reviewed in Han et al., 2013; Felice et al., 2016; Terunuma, 2018), and Sig1R has been established as a molecular target to treat the abovementioned disorders (Maurice and Su, 2009; Vavers et al., 2019). In addition, it has been demonstrated that the medial habenula plays an important role in stress, depression, memory and addiction (Viswanath et al., 2014). Functional GABA-B receptors are heterodimers whose function depends on the dimerization of the GABA-B R1 and GABA-B R2 subunits (reviewed in Bowery et al., 2002; Frangaj and Fan, 2018). It has been shown that mice lacking either the R1 or R2 subunit of the GABA-B receptor demonstrate altered anxiety and depression-related behavior (Mombereau et al., 2005), which is associated with behavioral changes found in C57Bl/6 background Sig1R−/− mice (Sabino et al., 2009; Chevallier et al., 2011; Couly et al., 2020). In addition, spontaneous seizures have been observed in GABA-B R2 subunit knockout mice (Mombereau et al., 2005). The medial habenula is divided into two subnuclei on the basis of cell type: cholinergic neurons are located in the ventral part of the medial habenula while Substance P-ergic neurons are located exclusively in the dorsal part of the medial habenula (Lee et al., 2019). By using Substance P and ChAT as markers of the dorsal and ventral parts of the medial habenula, respectively, we demonstrated that the decrease of the R2 subunit of the GABA-B receptor in Sig1R−/− mice is present in the ventral part of the medial habenula. In the brain, the medial habenula contains one of the highest concentrations of GABA-B receptors, suggesting the presence of strong inhibitory inputs (Viswanath et al., 2014). Presynaptic GABA-B receptors are abundant in cholinergic neurons of the medial habenula and control not only the release of acetylcholine but also glutamate (Lee et al., 2019). The reduced expression of the GABA-B receptor R2 subunit and disturbed GABA-B receptor function may explain the neurobehavioral phenotype of Sig1R−/− mouse and higher susceptibility to seizures.
GABA-B receptor agonists have been shown to diminish seizure activity in mouse models of both generalized convulsive and focal seizures (Joshi et al., 2016). Activation of GABA-B receptors with R-baclofen dose-dependently attenuated PTZ-induced kindling in CD-1 mice, while inhibition of GABA-B receptor by antagonists CGP 35348 and CGP 55845 resulted in more rapid development of kindling in the same model (De Sarro et al., 2000). It has been shown that activation of the GABA-B receptor in rat globus pallidus can reduce PTZ-induced tonic seizures (Chen et al., 2004). In patients with temporal lobe epilepsy significantly decreased number of GABA-B receptor binding sites were observed in the hippocampus (Furtinger et al., 2003). Taken together, these findings suggest that the reduced expression of the GABA-B receptor could explain why Sig1R−/− mice demonstrate a significantly increased susceptibility to PTZ- and BIC-induced tonic seizures. On the other hand, the GABA-B receptor agonist baclofen may possess a proconvulsant effect which is caused by a presynaptic GABA-B receptor-mediated inhibition of GABA release leading to disinhibition (Motalli et al., 1999; Han et al., 2013). Therefore, GABA-B receptor antagonists are thought to be promising for treating atypical absence seizures observed in Lennox-Gastaut syndrome (Han et al., 2013). In the medial habenula GABA-B receptors are known to mediate excitation through a unique signaling mechanism which comprises amplification of presynaptic Ca2+ entry through Cav2.3 channels and potentiation of co-release of glutamate and acetylcholine to excite interpeduncular neurons (Zhang et al., 2016). Interestingly, fenfluramine, a positive allosteric modulator of Sig1R (Martin et al., 2020), has been shown to be clinically effective in reducing seizures in patients with Lennox-Gastaut syndrome (Lagae et al., 2018), in which the most common seizure type is tonic seizures (Intusoma et al., 2013). Our data showed that Sig1R−/− mice demonstrate a significantly reduced threshold for tonic seizures, which suggests the possible role played by the Sig1R-GABA-B interaction in seizures in which the tonic component is the most common. Since GABA-B receptor-mediated mechanisms can be pro- and anti-convulsive (Joshi et al., 2016), this finding could explain the phenomena described for several Sig1R agonists and antagonists, which could also be pro- and anti-convulsive.
In the NE-100-induced seizure model, significantly reduced seizure severity was observed in Sig1R−/− animals and was presented as a lower seizure score, shorter duration and increased latency time of seizures compared to WT mice. These observations strengthen the pivotal role played by Sig1R in the control of seizures. However, the Sig1R antagonist NE-100 induced seizures not only in WT but also in Sig1R−/− animals. The mechanism of how NE-100 induces seizures is not known. We found that NE-100-induced seizures resulted in Sig1R-independent downregulation of iNOS and GABA-Aγ2 receptor subunit genes in both WT and Sig1R−/− animals, despite the different seizure severity observed in these animals after administration of NE-100. Recently, it was demonstrated that NE-100 can dose-dependently inhibit the exocytotic release of GABA and glutamate in synaptosomes isolated from rat brain (Pozdnyakova et al., 2020). In addition, bidirectional activity of NE-100 was observed on GABA uptake in rat brain synaptosomes (Pozdnyakova et al., 2020). Therefore, it seems that NE-100 dose-dependently disturbs the balance of excitation and inhibition. Since Sig1R−/− mice show decreased expression of the GABA-B R2 subunit and significantly lower seizure severity in the NE-100-induced seizure model, it is possible that the NE-100-induced seizure severity could be modulated through the level of the GABA-B R2 subunit and the activity of GABA-B receptor.
We also found that Sig1R−/− mice showed a decreased response to KCl and subsequently weaker maximal contraction in an isolated vasa deferentia model compared to WT animals. Recently, it was confirmed that Sig1R behaves as an atypical auxiliary subunit to modulate potassium channel function (Abraham et al., 2019). Therefore, the difference in the contractile response between WT and Sig1R−/− mice could be due to the different functions of potassium channels. It is known that activation of the postsynaptic GABA-B receptor activates inward-rectifier potassium channels (Bowery et al., 2002). The inhibitory action of Sig1R on the inward-rectifier potassium channel Kir2.1 has been previously demonstrated (Wong et al., 2016). Interestingly, the inhibition of inward-rectifier potassium channels by barium chloride results in an increase in contractions of isolated vasa deferentia(Diaz-Toledo and Jurkiewicz, 1991). In our study, the effect of barium chloride was less pronounced on the vasa deferentia isolated from Sig1R−/− mice, which confirms the role of inward-rectifier potassium channel function in this model. Similar to barium chloride, administration of NE-100 in an isolated vasa deferentia model resulted in a significant dose-dependent enhancement of electrical current-induced contraction amplitude in WT mice, while there was a significantly reduced effect of NE-100 on vasa deferentia isolated from Sig1R−/− mice. It should be noted that Sig1R agonists PRE-084, (+)-pentazocine and antagonists BD-1047 and NE-100 were shown to similarly inhibit the KCl-induced increase in [Ca2+]i in both WT and Sig1R−/− mouse brain synaptosomes and thus demonstrated Sig1R-independent effects of these ligands on KCl-induced depolarization (González et al., 2012). In keeping with previous findings, we also found a Sig1R-independent inhibitory effect of NE-100 on KCl-induced maximal contractility of WT and Sig1R−/− mouse vasa deferentia, which could be due to Sig1R-independent activity of NE-100 on inward-rectifier potassium channels.
5. Conclusions
Our results strongly demonstrate the reduced expression of the R2 subunit of the GABA-B receptor in the brain due to the genetic inactivation of Sig1R and supports the significance of Sig1R and GABA-B receptor cooperation in the modulation of the seizure threshold. Sig1R is a significant molecular target for seizure modulation and warrants further investigation for the development of novel anti-seizure drugs.