
Direct, gabapentin-insensitive interaction of a soluble form of the calcium channel subunit α2δ-1 with thrombospondin-4

By Ehab El-Awaad, Galyna Pryymachuk, Cora Fried, Jan Matthes, Jörg Isensee, Tim Hucho, Wolfram F. Neiss, Mats Paulsson, Stefan Herzig, Frank Zaucke, and Markus Pietsch
Excerpt from the article published in Scientific Reports 9, 16272 (2019), Published 07 November 2019 DOI: https://doi.org/10.1038/s41598-019-52655-y
Editor’s Highlights
- Thrombospondins (TSPs) form a family of five large oligomeric extracellular matrix glycoproteins that are expressed by numerous cell types.
- In neurons, astrocyte-secreted or recombinantly expressed TSP(s), particularly TSP-1, TSP-2 and TSP-4, were reported to promote the formation of excitatory synapses.
- The alpha2-detla (α2δ) proteins (α2δ‐1–4) are auxiliary subunits of voltage-gated calcium channels CaV1 and CaV2, and were found to be encoded by four different genes.
- The α2δ-1 protein is functionally involved in TSP-induced synaptogenesis. This interaction is crucial for excitatory synaptogenesis and neuronal sensitisation, suggesting a potential role in the pathogenesis of neuropathic pain.
- The interaction of dysregulated TSP-4 with α2δ-1 are partially independent of the role of the latter protein in regulating voltage-gated calcium channels’ trafficking and function.
- There is substantial evidence for a direct and specific Ca2+-insensitive TSP-4/α2δ-1 interaction which is rather weak.
Abstract
The α2δ‐1 subunit of voltage-gated calcium channels binds to gabapentin and pregabalin, mediating the analgesic action of these drugs against neuropathic pain. Extracellular matrix proteins from the thrombospondin (TSP) family have been identified as ligands of α2δ‐1 in the CNS. This interaction was found to be crucial for excitatory synaptogenesis and neuronal sensitisation which in turn can be inhibited by gabapentin, suggesting a potential role in the pathogenesis of neuropathic pain. Here, we provide information on the biochemical properties of the direct TSP/α2δ-1 interaction using an ELISA-style ligand binding assay. Our data reveal that full-length pentameric TSP-4, but neither TSP-5/COMP of the pentamer-forming subgroup B nor TSP-2 of the trimer-forming subgroup A directly interact with a soluble variant of α2δ-1 (α2δ-1S). Interestingly, this interaction is not inhibited by gabapentin on a molecular level and is not detectable on the surface of HEK293-EBNA cells over-expressing α2δ‐1 protein. These results provide biochemical evidence that supports a specific role of TSP-4 among the TSPs in mediating the binding to neuronal α2δ‐1 and suggest that gabapentin does not directly target TSP/α2δ-1 interaction to alleviate neuropathic pain.
Introduction
Thrombospondins (TSPs) form a family of five large oligomeric extracellular matrix glycoproteins that are expressed by numerous cell types, playing important roles in cellular migration, attachment and cytoskeletal dynamics1,2. Several TSP isoforms have been shown to be involved in a variety of physiological and pathological processes, including regulation of angiogenesis, apoptosis and platelet aggregation3,4,5. TSPs can be subdivided into subgroups A (TSP-1 and 2) and B (TSP-3–5, with TSP-5 also referred to as cartilage oligomeric matrix protein (COMP)) based on their oligomerisation state (trimeric or pentameric, respectively) and domain structure (Fig. 1A). In neurons, astrocyte-secreted or recombinantly expressed TSP(s), particularly TSP-1, TSP-2 and TSP-4, were reported to promote the formation of excitatory synapses both in vitro and in vivo through interaction with the voltage-gated calcium channel subunit α2δ-16,7,8,9,10. The α2δ proteins (α2δ‐1–4) are auxiliary subunits of voltage-gated calcium channels CaV1 and CaV2, and were found to be encoded by four different genes11,12,13. Functions of these auxiliary subunits include the modulation of trafficking, expression in the plasma membrane14,15,16,17, and biophysical properties of the channels15,17,18,19. Importantly, α2δ-1 acts as a specific binding site for gabapentinoid drugs20,21, mediating their analgesic effect in neuropathic pain21,22,23. Furthermore, studies using different animal models of neuropathic pain indicated the involvement of α2δ-1 in pain development, with nerve injuries leading to up-regulation of α2δ‐1 in both dorsal root ganglion (DRGs) and spinal dorsal horn neurons24,25,26,27 as well as to an increase of miniature excitatory post-synaptic current (mEPSC) frequency in the latter neurons22,25,26,28,29,30. Similarly, injury-induced TSP-4 is reported to mediate central sensitisation and neuropathic pain states8,9,31,32,33,34,35,36,37. This effect was recently shown to be mediated by activation of a TSP-4/α2δ‐1-dependent pathway which requires a direct molecular interaction between the two proteins9,34. Furthermore, the presence of TSP-4 was shown to modestly but significantly reduce the binding affinity of 3H-gabapentin (3H-GBP) towards α2δ‐1 in membrane preparations from TSP-4/α2δ‐1 co-transfected cells38. Taken together, the TSP-4/α2δ‐1 protein-protein interaction seems to be of potential translational importance and thus may serve as a novel target for developing a new class of analgesics against neuropathic pain.
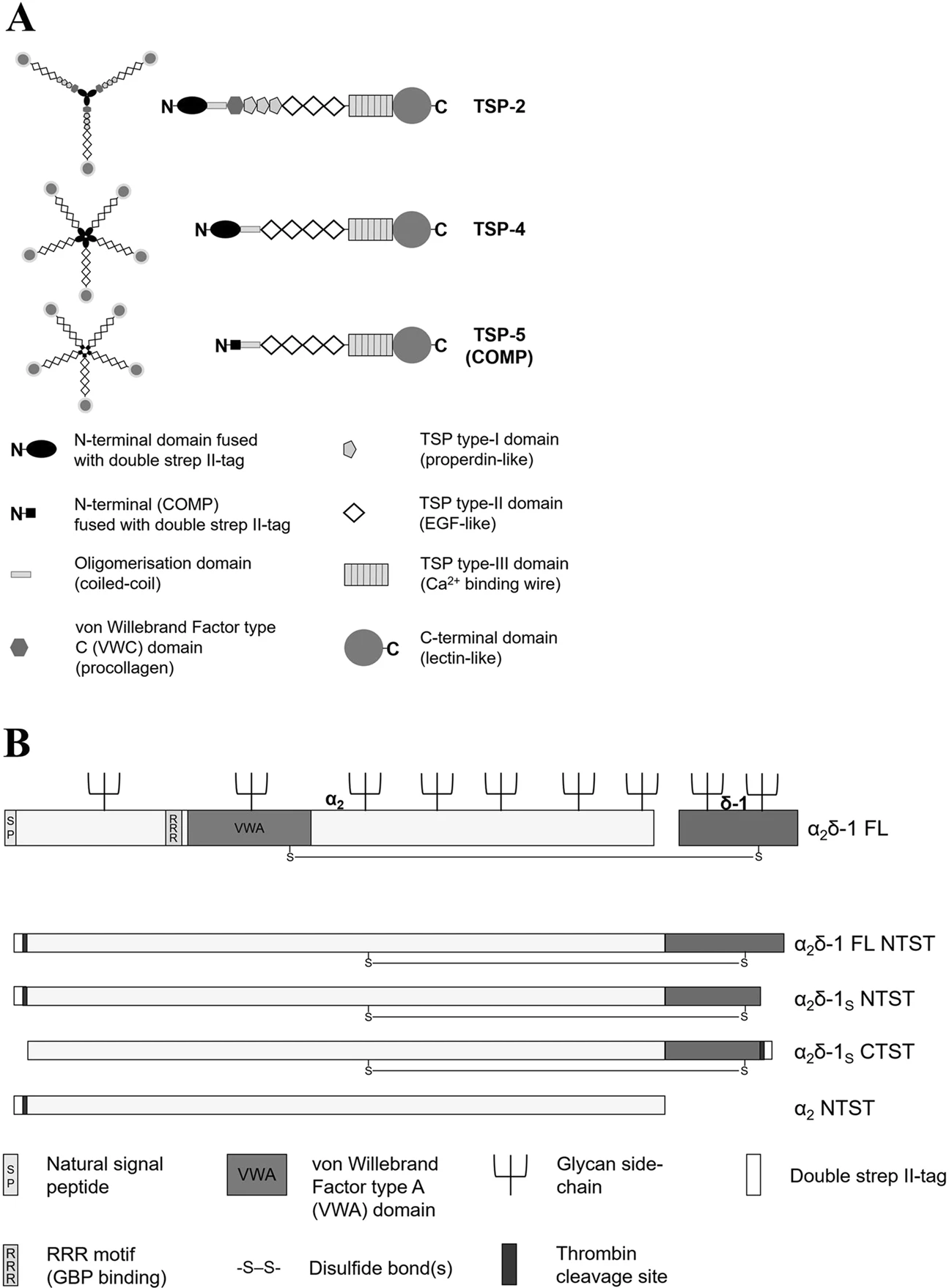
Schematic presentation of the structures of the recombinant proteins generated in this study. (A) Domain structure and oligomerisation state of the generated recombinant full-length TSP-2 (trimer), TSP-4 and COMP (pentamers). Schematic representation adapted by permission from Springer Nature, Cell Mol Life Sci, Structures of thrombospondins, Carlson, C. B., Lawler, J. & Mosher, D. F., Copyright (2008)39. All recombinant TSPs have been expressed with an N-terminal double strep II-tag and contain glycan side-chains which are not shown for reasons of clarity. (B) Structure of α2δ-1 FL protein (adapted from Cell 139, Eroglu, Ç. et al., Gabapentin receptor αδ2δ-1 is a neuronal thrombospondin receptor responsible for excitatory CNS synaptogenesis, 380–392, Copyright (2009), with permission from Elsevier7) and simplified depiction of the derived non-proteolytically processed α2δ-1 mutants generated in this study. The RRR motif, the von Willebrand Factor type A domain, and the glycan side-chains are not shown in the α2δ-1 mutants for reasons of clarity.
The aim of the present study is to investigate the biochemical characteristics of the direct molecular interaction between TSPs and α2δ‐1, addressing the question whether α2δ‐1 binding is specific to TSP-4 or redundant among other TSPs. GBP has been shown so far to inhibit the interaction of α2δ‐1 with a truncated form of TSP-2 in co-immunoprecipitation experiments7 as well as functionally by inhibiting synaptogenesis7,8,10, neuron sensitisation and behavioural hypersensitivity induced by TSP-2, its truncated fragment and/or TSP-49,34,35. Thus, we examined whether the direct TSP/α2δ‐1 interaction can be inhibited by GBP on a molecular level as well. We therefore generated purified recombinant forms of three full-length TSPs (TSP-2, TSP-4 and COMP) as well as soluble forms of α2δ‐1 subunit (α2δ‐1S), that shows GBP binding affinity similar to that of wild-type α2δ-1, and the α2 peptide chain of α2δ‐1 (Fig. 1). Both the interaction of these recombinant TSPs with α2δ‐1 and the possible inhibition by GBP were examined in a solid-phase ELISA-style ligand binding assay, with the capability of soluble α2δ‐1 to interact with GBP being proven by a newly developed surface plasmon resonance (SPR)-based binding assay. In order to demonstrate the characteristics of the direct TSP-4/α2δ‐1 interaction in an environment similar to that of native cells, we attempted to visualise the interaction of fluorescently labelled TSP-4 with membrane-localised full-length (FL) α2δ‐1 in a cell-based system.
Results
Biochemical characteristics of recombinant purified proteins expressed in HEK293-EBNA cells
To investigate the direct binding of different TSPs to α2δ-1, three full-length recombinant TSPs, namely, the trimeric TSP-2, the pentameric proteins TSP-4 and COMP, and a soluble C-terminal deletion mutant of α2δ-1 carrying an N-terminal (α2δ-1S NTST) or a C-terminal (α2δ-1S CTST) double strep II-tag (Fig. 1) were generated in a eukaryotic expression system. Coomassie stained gels and immunoblots of the purified proteins confirmed their purity, identity and integrity (Fig. 2; Supplementary Fig. S1). As expected for TSPs, the intact proteins showed high molecular weight bands with approximate apparent molecular weights (Mr) in the range ~500–670 kDa when compared to laminin-111 and thyroglobulin as marker proteins (Fig. 2A). The oligomerisation patterns of these high molecular weight TSPs (i.e. pentamers for TSP-4 and COMP and trimer for TSP-2) were confirmed by comparing immunoblots in the absence and presence of the reducing reagent DTT (Fig. 2B; Supplementary Fig. S1a). In the latter case, DTT reduces the interchain disulphide bonds within the oligomerisation domains of the analysed TSPs39 and major bands of monomeric proteins with Mr of TSP-2 (~240 kDa), TSP-4 (~160 kDa), and COMP (~130 kDa) were observed (Fig. 2B, +DTT; Supplementary Fig. S1a, +DTT).
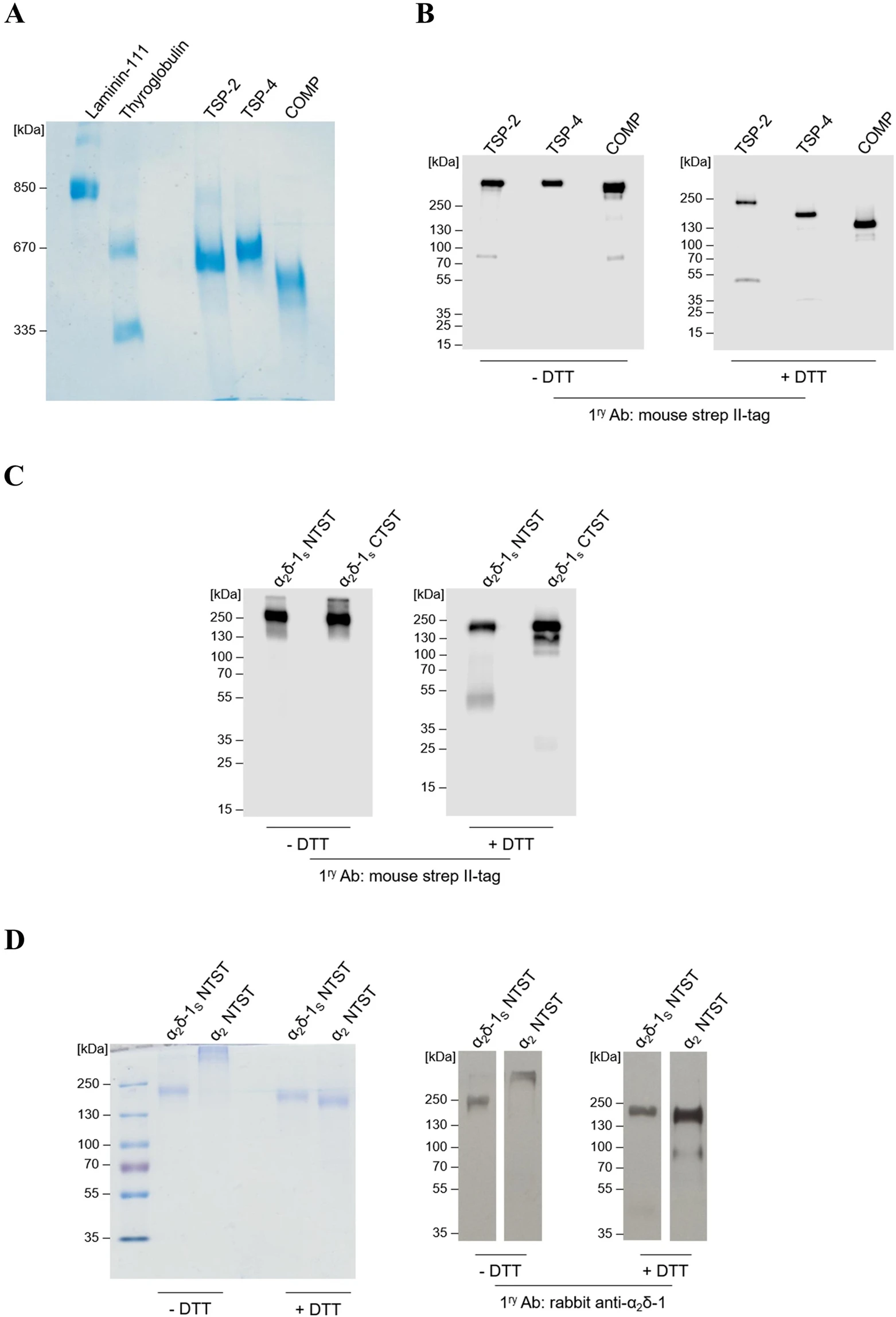
The generated recombinant TSPs and α2δ-1S variants show high degree of purity and integrity in Coomassie staining and western blot analyses. (A,D left) Representative Coomassie-stained gels and (B,C and D right) immunoblots of three full-length TSP proteins, all carrying an N-terminal double strep II-tag: TSP-2, TSP-4, and COMP (A,B); α2δ-1S variants carrying either an N-terminal (α2δ-1S NTST) or a C-terminal (α2δ-1S CTST) double strep II-tag (C); α2δ-1S NTST and α2 peptide chain carrying an N-terminal double strep II-tag, α2 NTST (D). Proteins were separated under non-reducing (−DTT) or reducing conditions (+DTT) on 4–15% (B), 10% (C), and 7% (D) polyacrylamide gels, respectively, while in (A) proteins were separated on 0.5% agarose (w/v)/3% polyacrylamide (w/v) composite gels without prior DTT treatment. Proteins were either stained with colloidal Coomassie stain (A,D left) or detected with the following primary antibodies after blotting: mouse anti-strep II-tag (B,C) or rabbit anti-α2δ-1 (D right). Secondary antibodies included the polyclonal rabbit anti-mouse IgG (B,C) and swine anti-rabbit IgG (D right), both conjugated with horseradish peroxidase (see Supplementary Table S2 for further information). In all gels the molecular weight standard (in kDa) indicated on the left was PageRuler Plus Prestained Protein Ladder (Thermo Fisher Scientific) except for (A) in which both thyroglobulin (Sigma) and recombinant laminin-111 (kind gift from Prof. Dr. Monique Aumailley, Institute for Biochemistry II, Centre for Biochemistry, Medical Faculty, University of Cologne) were used.
Similar analysis was performed for the generated α2δ-1S NTST and the respective C-terminally tagged α2δ-1 variant, α2δ-1S CTST, showing single but smeared bands at approximate Mr ~200 kDa under non-reducing conditions (Fig. 2C, −DTT; Supplementary Fig. S1b, −DTT) and appear as distinct bands at approximate Mr ~180 kDa under reducing conditions (Fig. 2C, +DTT; Supplementary Fig. S1b, +DTT). So far, there is no comprehensive explanation for this gel band shift in the presence of DTT. However, it cannot be attributed to the reductive cleavage of the interchain disulphide bridge between α2 and δ-1 followed by loss of the smaller δ-1 chain. This conclusion arises from the observation that α2δ-1S bearing the C-terminal double strep II-tag (α2δ-1S CTST) is still detectable in immunoblots probed with strep II-tag antibody following DTT treatment. In agreement with this result, mass spectra of α2δ-1S NTST recorded with and without DTT pre-treatment showed almost identical molecular ion peaks ([M + H]+; Table 1, Supplementary Fig. S2d,e). This observation confirms the results by Brown and Gee40 who first described a similar soluble mutant of the porcine α2δ-1 orthologue which retains high affinity for 3H-GBP. Although uncleaved α2δ-1 may not represent a functional form as a subunit of the CaV channels and can inhibit native calcium currents in mammalian neurons41, the TSP/α2δ-1 pathway is thought to be at least partially independent of the roles of α2δ-1 as a CaV channel subunit7,10. Therefore, the recombinant uncleaved α2δ-1S variant used in this study should be suitable for the purpose of investigating TSP binding biochemically. Notably, we observed a minor band in the immunoblots of α2δ-1S CTST at Mr ~25 kDa upon DTT treatment and detection with strep II-tag antibody (Fig. 2C, +DTT) which is most likely attributed to cleaved strep-tagged δ-1Schain. This indicates the presence of a small fraction of the generated purified α2δ-1S in a proteolytically cleaved form.
| Protein | m/zcalc [M + H]+ | m/zexp [M + H]+ |
|---|---|---|
| TSP-2 (monomer) | 132,137 | 146,958 |
| TSP-4 (monomer) | 107,409 | 109,351 |
| COMP (monomer) | 84,714 | 88,463; 87,178 |
| α2δ-1S NTST (−DTT) | 123,000 | 152,236 |
| α2δ-1S NTST (+DTT) | 112,130* | 152,258 |
The m/zcalc [M + H]+ values for all proteins (in Da) were calculated based on their amino acid sequences using ExPASy Compute pI/Mw online tool, while the m/zexp [M + H]+ values (in Da) were obtained from the MALDI-TOF MS spectra of the respective proteins.
* m/zcalc [M + H]+ of α2 NTST was calculated for the expected product of the DTT-mediated reduction of the disulphide bond between α2 and δ-1S in α2δ-1S NTST.
In addition to the α2δ-1S variants generated, the α2 peptide chain (α2 NTST) was recombinantly produced in a similar way (Fig. 1B). Expression and purification of this fragment as well as analyses by SDS-PAGE and immunoblotting (Fig. 2D) were carried out as described above. Here, we observed the formation of a high molecular weight product under non-reducing conditions which dissociated into the monomeric form after DTT treatment (approximate Mr ~ 170 kDa, Fig. 2D) which points to the formation of interchain disulphide bonds in the absence of reducing agents (see also Discussion section below).
Notably, all recombinant proteins generated in this study that had been analysed by SDS-PAGE and Western Blot showed protein bands at remarkably higher Mr than expected from their amino acid sequences. It is known that the electrophoretic mobility of proteins can be greatly influenced by the extent of post-translational modifications (e.g. glycosylation) of the protein where the glycan chains do not bind SDS leading in many cases to decreased mobility, and increased Mr, of the glycoprotein analysed by SDS-PAGE42. In addition, sample treatment prior to loading onto the gel (e.g. heating at 95 °C with DTT) represents a possible source of abnormal protein migration on the gels through its impact on the structure of the analysed protein43. Therefore, further analysis was carried out to determine the accurate molecular masses of the recombinant purified proteins using MALDI-TOF mass spectrometry. The results are shown in Table 1 and Supplementary Fig. S2. The molecular masses of TSP-4 and COMP, both in monomeric form, were found to be only slightly higher (1–3 kDa) than the theoretical masses calculated on the basis of each protein’s amino acid sequence, indicating minor post-translational modifications, e.g. glycosylation, of these proteins. In case of TSP-2 and α2δ-1S NTST, the experimentally determined masses were about 15 and 30 kDa, respectively, larger than the theoretical ones, which is likely attributed to heavy glycosylation, as shown previously for α2δ-1 in the work of Kadurin et al.44 (Table 1, Supplementary Fig. S2a,d,e).
Characterisation of the interaction of TSPs with α2δ-1S using an ELISA-style ligand binding assay
First, the purified recombinant full-length TSPs (soluble) were examined for their direct interaction with immobilised α2δ-1S NTST in an ELISA-style ligand binding assay, that was validated using the model interaction of COMP and matrilin-3 proteins45 (Supplementary Fig. S4). Preliminary experiments with TSP concentrations up to ~500 nM had demonstrated that out of the three TSPs generated in this study, only TSP-4 was able to directly interact with α2δ-1S NTST (data not shown). To rule out the possibility of a very low binding affinity of TSP-2 and COMP towards α2δ-1, we finally utilised a comparably high concentration (1,000 nM) of all TSP proteins in the same ELISA which confirmed our preliminary data (Fig. 3A). Therefore, we decided to focus on the interaction of TSP-4 with α2δ-1S and study it more in detail. Notably, we detected comparable binding signals of TSP-4 to either α2δ-1S NTST or α2δ-1S CTST variants in a preliminary experiment (data not shown) and hence we chose to proceed with one of the two variants, namely, α2δ-1S NTST in our binding assays. Titration of immobilised α2δ-1S NTST with increasing concentrations of TSP-4 (11-1,505 nM) showed saturable binding with an apparent KD value of about 200 nM. Since Ca2+ binding is associated with major conformational changes and structural rearrangements of both TSPs46,47 and the metal ion-dependent adhesion site (MIDAS) motif of α2δ-117,48, we investigated the effect of Ca2+ on TSP-4/α2δ-1S NTST interaction. Our data show that the apparent KD value was slightly, but not significantly, decreased in presence of 1 mM Ca2+(Table 2, Fig. 3B). Similarly, the maximum binding value (Bmax, unitless) showed a small, statistically non-significant increase in presence of 1 mM Ca2+ (Table 2, Fig. 3B). These results indicate that the TSP-4/α2δ-1S NTST interaction is not sensitive to Ca2+ changes. Next, the ability of TSP-4 to directly interact with the α2 fragment of α2δ-1 was analysed. Here, we observed a two-fold, statistically significant increase in the binding signal of a single concentration of TSP-4 (1,000 nM) when using immobilised α2 NTST instead of α2δ-1S NTST (Fig. 3C). These results indicate the localisation of the TSP-4 binding site(s) within the α2region of α2δ-1 in agreement with data obtained by Eroglu et al.7. The enhanced binding signal of TSP-4 to α2 NTST is suggestive of a more favourable conformation of α2 NTST which is more accessible for TSP-4 binding, as compared to the non-proteolytically cleaved α2δ-1S NTST.
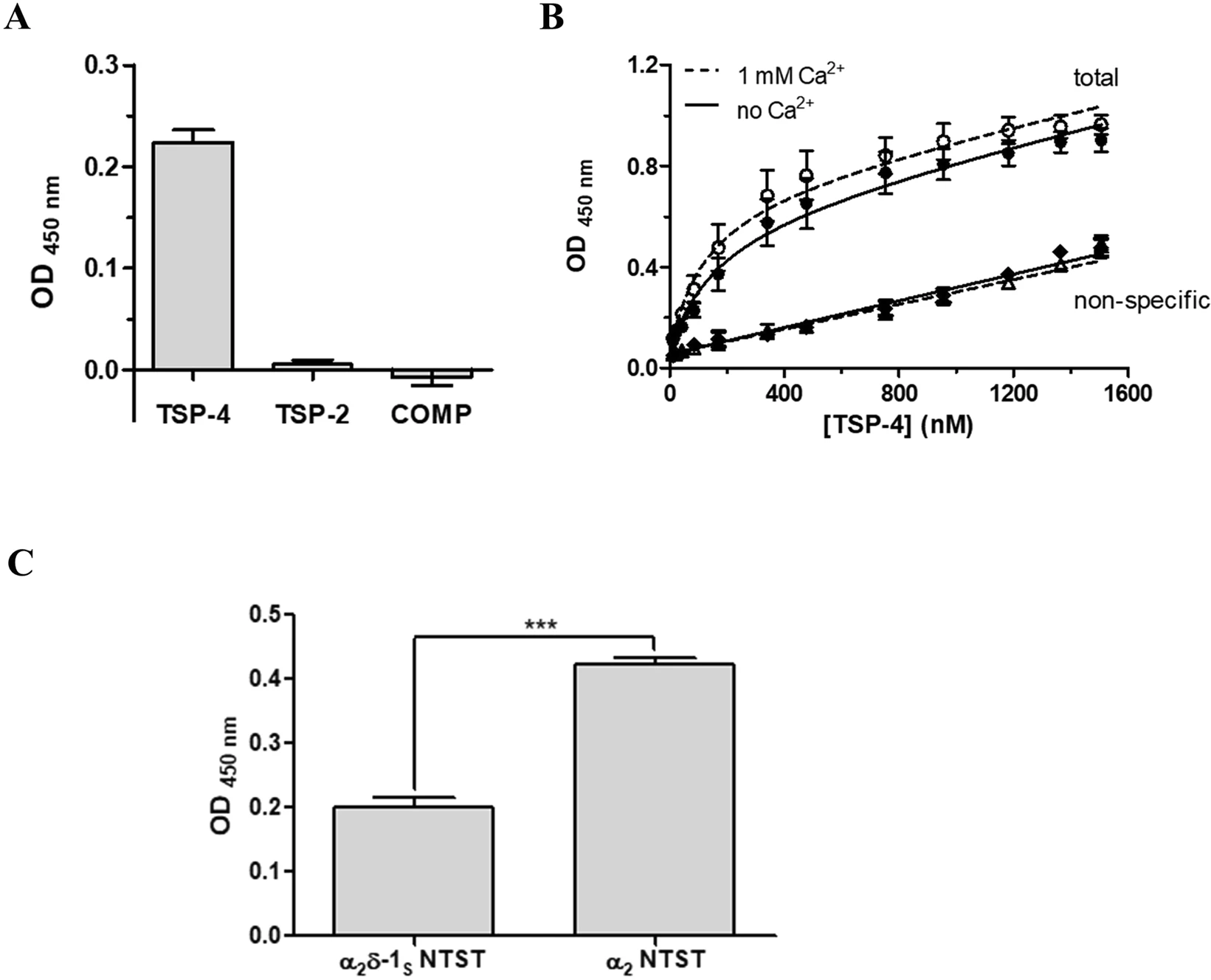
The direct binding to α2δ-1S NTST is TSP-4 specific in an ELISA-style ligand binding assay. α2δ-1S NTST (20 µg/ml) was coated onto 96-well plates and incubated with either (A) TSP-2, TSP-4 or COMP (1,000 nM), or (B) increasing concentrations of TSP-4 (11–1,505 nM). Shown are data for total (circles) and non-specific (triangles, rhombi) binding in the absence (full symbols) and presence (open symbols) of 1 mM Ca2+. (C) Soluble α2 NTST or α2δ-1S NTST (10 µg/ml) were coated onto 96-well plates and incubated with TSP-4 (1,000 nM). Binding assays (A,C) were carried out in the presence of 1 mM Ca2+ and bound proteins were detected with the corresponding TSP-specific antibody/antiserum (see Supplementary Table S2). Specific binding in (A,C) was calculated by subtracting OD values of non-specific binding from those of total binding. Data of total and non-specific binding were used to calculate KDand Bmax in (B), with the linear dependence of the non-specific signal on the TSP-4 concentration ensuring the absence of perturbations of the assay system. Data represent mean values ± SEM of 3 independent measurements performed in duplicates or triplicates. Statistical analysis in (C) was done using unpaired two-tailed Student’s t-test (***P = 0.0003).
| Assay buffer | KD (nM) | Bmax |
|---|---|---|
| TBS | 198 ± 49 | 0.579 ± 0.066 |
| TBS + 1 mM Ca2+ | 153 ± 46 | 0.677 ± 0.054 |
Data represent mean values ± SEM of 3 independent experiments, each performed in duplicate. Statistical analysis by an unpaired two-tailed Student’s t-test showed no significant difference for KD (apparent dissociation constant, P = 0.5381) and Bmax (maximum binding, unitless, P = 0.3138) obtained in the absence and presence of Ca2+.
To investigate the effect of the known α2δ-1 ligand GBP20 on the observed TSP-4/α2δ-1Sinteraction, we checked for the ability of the recombinantly expressed mutant α2δ-1S NTST to bind GBP with high affinity (KD = 219 nM) using a label-free surface plasmon resonance (SPR) assay (Fig. 4A, Supplementary Fig. S3). Our results are in agreement with the binding data for 3H-GBP to membrane preparations of heterologously expressed human α2δ-1 (KD range of 140–175 nM)38. Various GBP concentrations (0.05–1,000 µM) were used in the TSP-4 /α2δ-1SNTST binding assay to interfere with this protein-protein interaction but did not show inhibitory effects (Fig. 4B). To rule out the possibility that the GBP binding pocket might not be accessible after immobilisation of α2δ-1S NTST on the ELISA microplate, we had coated the wells with α2δ-1S NTST after pre-incubating the protein with GBP. In addition, GBP had been supplemented to the liquid phase during blocking and incubation with TSP-4 ensuring availability of a sufficient number of GBP molecules to α2δ-1S NTST and preventing dissociation of bound GBP during the experiment. In a further experiment, the binding of various concentrations of TSP-4 (12.5–1,000 nM) to α2δ-1S NTST was not affected by the highest GBP concentration (1,000 µM) investigated (Fig. 4C). Together, these data suggest that GBP alone is not sufficient to disrupt the interaction of TSP-4 with α2δ-1 on a molecular level.
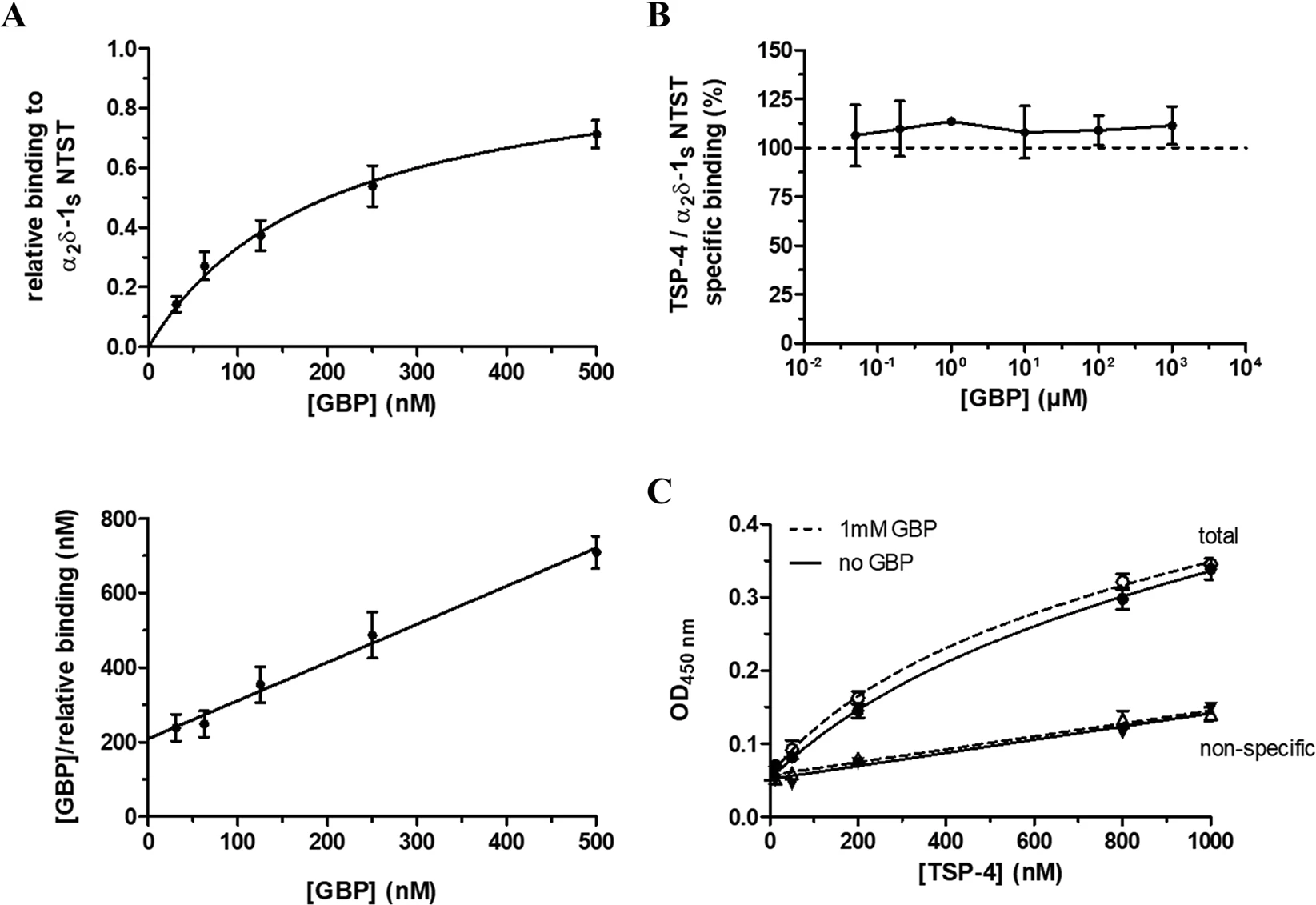
GBP does not directly interfere with the binding of TSP-4 to α2δ-1S NTST in an ELISA-style ligand binding assay. (A) Surface plasmon resonance (SPR) measurements of the binding of GBP to recombinant α2δ-1S NTST. The protein (10–15 µg/ml) was directly immobilised to CM5 sensor chips and GBP (31.25–500 nM) in PBS buffer, pH 7.4 containing 0.05% Tween 20 was passed over the chip at a flow rate of 30 μl/min. Shown are data of the relative GBP binding to α2δ-1S NTST (Top) obtained from single cycle kinetics protocol (mean values ± SEM of 4 independent experiments, Fig. S3). Each experiment was analysed by non-linear regression according to the equation RU/RUmax = [GBP]/(KD + [GBP]), where the ratio of the binding response and the maximum binding response, RU/RUmax, represents the relative binding at a given GBP concentration, [GBP], and KD is the dissociation constant of the two interaction partners. Data analysis yielded a value of KD = 219 ± 47 nM (mean value ± SEM, n = 4), with the linear shape of the Hanes-Woolf transformation (Bottom) showing equimolar binding of the two interaction partners. For the ELISA-style assay, the α2δ-1S NTST (10 µg/ml) protein was coated onto 96-well plates and incubated with either (B) TSP-4 (1,000 nM) in the absence and presence of increasing concentrations of GBP (0.05–1,000 µM), or (C) increasing concentrations of TSP-4 (12.5–1,000 nM) in the absence (full symbols) and presence (open symbols) of GBP (1,000 µM). The assay was carried out in the presence of 2 mM Mg2+ and bound TSP-4 was detected with TSP-4-specific antiserum. Specific binding was calculated by subtracting OD values of non-specific binding (triangles) from those of total binding (circles). Data in (B) and (C) represent mean values ± SEM of 2 to 3 independent experiments, each performed in duplicate. In (B) the OD values for specific binding of TSP-4 in the presence of GBP (0.05–1,000 µM) were normalised to those in the absence of GBP.
Fluorescent A555-TSP-4 does not bind to membrane-bound α2δ-1 in a cell-based binding assay
To examine the interaction between TSP-4 and full-length membrane-bound α2δ-1 in a cellular environment, HEK293-EBNA cells were transfected with either empty vector or vector encoding α2δ-1 FL NTST. Immunocytochemical analysis of cells transfected with α2δ-1 FL NTST revealed a marked increase in α2δ-1 immunoreactivity, confirming the over-expression of heterologous α2δ-1 (Fig. 5A, left column).
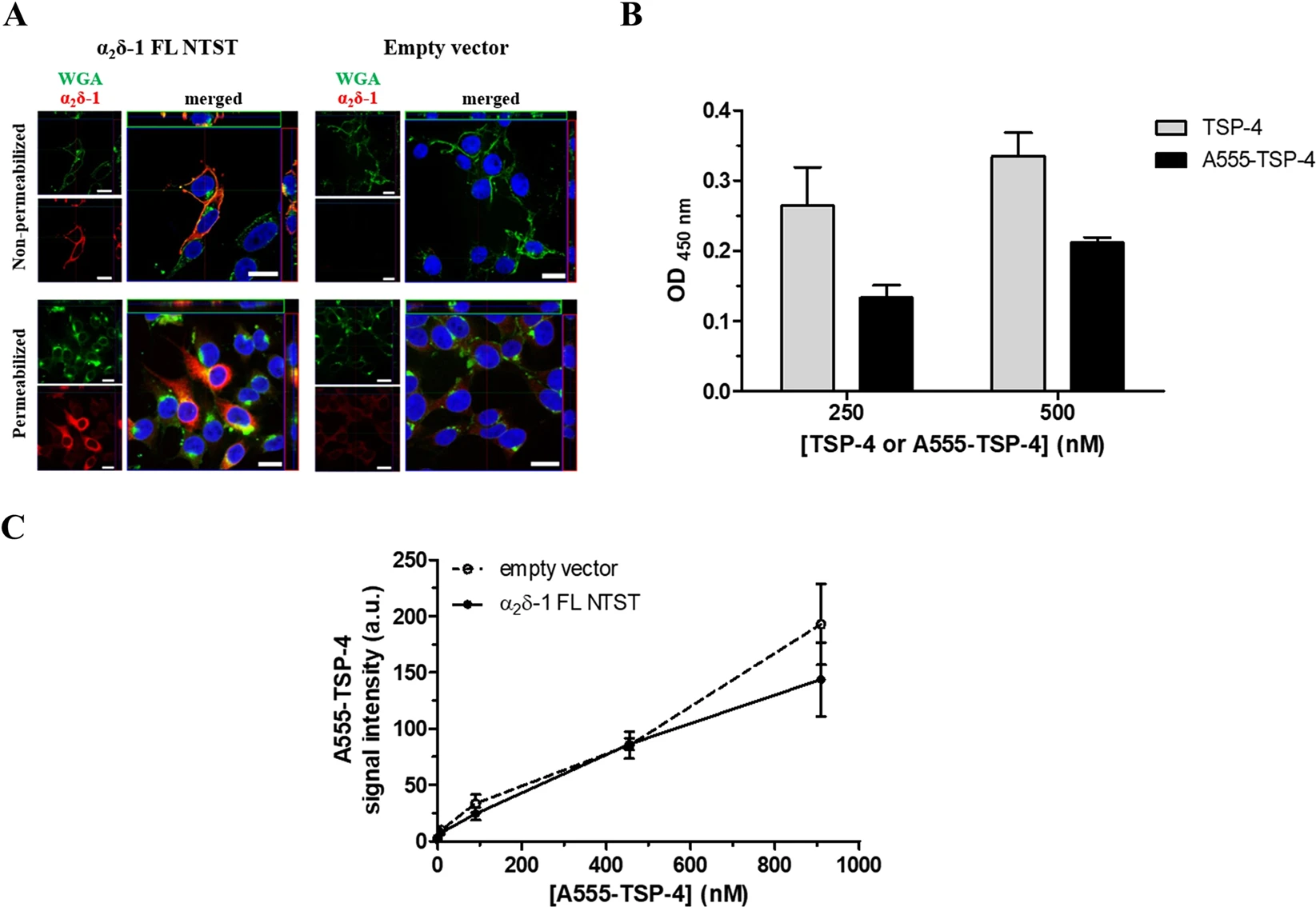
Fluorescent A555-TSP-4 protein does not bind to membrane-bound α2δ-1 in HEK293-EBNA cells. (A) Representative confocal images of the immunocytochemical detection of membrane-localised α2δ-1 FL NTST over-expressed in HEK293-EBNA cells. Cells were transfected with either empty vector (right column) or vector encoding full-length α2δ-1 containing an N-terminal strep II-tag (α2δ-1 FL NTST, left column). Cells were stained either without permeabilisation (upper row) or after membrane permeabilisation (lower row). Signals of WGA conjugated with Alexa Fluor 633 (green) and α2δ-1 (red) are shown individually in the small images; merged signals are shown in the large images. DAPI was used to visualise the nucleus (blue). Images show top view as well as upper-side (green box) and right-side (red box) views of a single slice of scanning near the middle of cells. Scale bar is 20 µm for all images. (B) Binding of A555-TSP-4 or TSP-4 to α2δ-1S NTST analysed by an ELISA-style ligand binding assay. α2δ-1S NTST (20 µg/ml) was coated onto 96-well plates and incubated with either TSP-4 or A555-TSP-4 at two different concentrations (250 and 500 nM) for each protein. Specific binding was calculated by subtracting OD values of non-specific binding from those of total binding. Data represent mean ± SEM of 2 independent measurements, each performed in duplicate. OD values for specific binding of TSP-4 and A555-TSP-4 were subjected to unpaired Student’s t-test. No significant difference was found (P > 0.05) for each of the two concentrations of both TSP-4 species used. (C) Cells transfected with either empty vector or vector encoding α2δ-1 FL NTST were incubated with increasing concentrations of A555-TSP-4 (final concentration: 9–909 nM) in 96-well imaging plates. Control wells received the same volume of dilution medium (40 µl) without A555-TSP-4. Average A555-TSP-4 fluorescent signal intensities from wells containing cells transfected with α2δ-1 FL NTST encoding vector or empty vector (±SEM of 3 independent experiments, each performed in triplicate) were plotted versus the A555-TSP-4 concentration.
In cells immunostained without permeabilisation, a high degree of co-localisation of α2δ-1 and wheat germ agglutinin (WGA) used for labelling glycoproteins or glycolipids of the outer leaflet of the plasma membrane49 were observed, assuring the localisation of heterologous α2δ-1 in the plasma membrane (Fig. 5A, upper left image). The transfected cells were incubated with increasing concentrations of fluorescently labelled A555-TSP-4 (9–909 nM), the binding of which to α2δ-1S NTST was found to be non-significantly lower than that of unlabelled TSP-4 in ELISA-style assay (Fig. 5B). Images of the stained cells were acquired using a High Content Screening microscope and the average A555-TSP-4 signal intensity was determined for each well. In addition, α2δ-1 FL NTST was visualised by immunostaining to analyse the co-localisation of A555-TSP-4 with α2δ-1 FL NTST. Results show low overall A555-TSP-4 signal intensity which increases upon increasing the concentrations of the fluorescent protein. However, no difference was observed when comparing α2δ-1 FL NTST- and empty vector-transfected cells (Fig. 5C). Furthermore, no co-localisation of A555-TSP-4 and α2δ-1 FL NTST signals was detected in these experiments (Supplementary Fig. S5). These results are in accordance with recently published data by Lana et al.38 showing no TSP-4 co-localisation with α2δ-1 on the cell surface of tsA-201 cells co-expressing both proteins or in mixed populations of cells transfected separately with either α2δ-1 or TSP-4.
Discussion
All TSPs, i.e. TSPs 1–4 and COMP, were previously identified as synaptogenic proteins which, together with other astrocyte-derived factors, help to promote the formation of functional excitatory synapses in the CNS6,7. The α2δ-1 protein was demonstrated to be functionally involved in TSP-induced synaptogenesis by means of synaptic assays in retinal ganglion cells (RGCs)7, DRG/spinal cord primary neuron co-culture8,9, purified cortical neurons10 as well as in dorsal spinal cord of mice34. Biochemically, α2δ-1 was reported to interact in co-immunoprecipitation experiments with TSP-1, TSP-2 and TSP-4 from rat cerebral cortex7 as well as with TSP-4 from rodent spinal cord34. Similarly, a TSP-2 fragment containing all three EGF-like repeats, the calcium-binding repeats, and the C-terminal globular domain was co-purified with full-length α2δ-1 or its protein-binding VWA domain after co-expression in HEK293 cells7. Recently, Park et al.9,34 demonstrated for the first time a direct molecular interaction between α2δ-1 and recombinant full-length TSP-4 or its fragments containing EGF-like or coiled coil domains. In the present study, we investigated the biochemical properties of the direct TSP-4/α2δ-1 interaction. Furthermore, it was of importance to know whether other members of the TSP protein family are also able to directly bind to α2δ-1 in an analogous manner to that of TSP-4. Our data demonstrated that only full-length TSP-4, but not TSP-2 or COMP, is able to directly interact with immobilised soluble α2δ-1 variant (α2δ-1SNTST) in an ELISA-style ligand binding assay (Fig. 3A), indicating the specificity of this protein-protein interaction. This observation is in direct contrast to that of Eroglu et al.7 (see above). Nevertheless, TSP-4 is remarkably the only isoform of TSP proteins reported so far to be implicated in neuropathic and joint-mediated chronic pain in rodents along with neuronal α2δ-18,9,31,34,35. During the processes resulting in such pain, both TSP-4 and α2δ-1 are up-regulated on the protein level and temporally correlate with the development of behavioural hypersensitivity in the respective animal models (for review see ref.50). In contrast, TSP-1/-2, though previously shown to be up-regulated after ischemic brain injury in rodents4,51 and promoting the subsequent synaptic recovery51, are not dysregulated on the protein level in dorsal spinal cord after spinal nerve ligation in mice, even when behavioural hypersensitivity was evident in these animals. This observation led Kim et al.31 to rule out the possibility of the involvement of these two astrocyte-secreted proteins in mediating TSP/α2δ-1-induced neuropathic pain. With regards to COMP, it has been reported to be expressed in several tissues including skeletal muscle, tendon, and cartilage. In the latter tissue, COMP is known to be mainly involved in chondrocyte differentiation, attachment, and cartilage extracellular matrix assembly52,53,54,55, with mutations in COMP being associated with pseudoachondroplasia56,57. COMP in skeletal muscle, tendons, and perichondrium can be theoretically in contact with nerve terminals containing α2δ-1. In addition, COMP was shown to have synaptogenic potential in RGCs (as discussed) and shares a high degree of both overall sequence identity (~70%) and structural similarity (Fig. 1A) with TSP-4. That is why we investigated COMP as a potential interaction partner of α2δ-1 in our binding studies. Nevertheless, COMP is, in contrast to TSP-4, neither abundant in neurons and astrocytes nor it is known to be dysregulated in neuropathic pain states. The fact that, beside TSP-4, all other TSPs were previously found to induce synaptogenesis through a mechanism involving neuronal α2δ-17 may refer to other cellular factors or scaffold proteins required to mediate their interaction with α2δ-1 indirectly. Furthermore, it is tempting to speculate that TSP-induced synaptogenesis might be of little relevance to neuropathic pain development since, as previously mentioned, TSP-4 is the only member of TSP family found to be up-regulated in dorsal spinal cord following nerve injury. Indeed, a recent study shows that an enhanced presynaptic NMDA receptor activity, rather than synaptogenesis, is responsible for maintaining the increased synaptic excitatory transmission in dorsal spinal cord leading to chronic pain states following nerve injury in mice58.
In further experiments, we observed a significantly increased binding of TSP-4 to α2 NTST when compared to α2δ-1S NTST (Fig. 3C), confirming previous data by Eroglu et al.7demonstrating the TSP-4 binding site to be localised within the α2 region of α2δ-1 (VWA domain). The observed enhancement in binding towards α2 might be attributed to more exposed TSP-4 binding motif(s) in the immobilised α2 NTST compared to the non-proteolytically processed α2δ-1S NTST. This result is in agreement with recent findings from Lana et al.38 where wild-type α2δ-1 was very weakly co-immunoprecipitated with TSP-4, but no co-immunoprecipitation of the mutant α2δ-1 (MIDASAAA) with TSP-4 was detected in lysates of co-transfected tsA-201 cells. In addition, the binding to α2 NTST seems again to be TSP-4-specific since negligible binding signals were detected when equimolar concentrations of either COMP or a truncated TSP-4 fragment were utilized in a pilot experiment (data not shown). Although our results are supported by reported data, we cannot rule out the possibility that the enhanced TSP-4 binding signal is due to improperly expressed α2 NTST since unpaired cysteine residues, normally involved in the formation of disulphide bridges with other cysteine residues in the deleted regions of wild-type α2δ-159, become available. It is worth mentioning here that a very high tendency for multimerisation was observed for a recombinant VWA domain of α2δ-1 generated in this study (data not shown) due to the formation of intermolecular disulphide bonds. Therefore, it might be appropriate to consider the expression of α2 and VWA fragments in which the unpaired cysteines are replaced by other isosteric residues (e.g. serine) for future binding studies.
One of the reported small molecules capable of interfering with the TSP/α2δ-1 interaction is GBP, an approved analgesic against neuropathic pain60,61,62 and a known ligand of α2δ-120. Biochemically, co-immunoprecipitation experiments showed that the interaction between a truncated TSP-2 fragment and α2δ-1 FLAG in a co-culture of two populations of HEK293 cells was diminished in the presence of GBP7. In addition, as previously mentioned, TSP-4 modestly but significantly reduces the binding affinity of 3H-GBP to α2δ-1, suggesting rather an allosteric than a pure competitive mode of inhibition38. Furthermore, in vivo data revealed the ability of GBP to block TSP-4-induced neuronal sensitisation and behavioural hypersensitivity as well as changes in Ca2+ currents and intracellular Ca2+ transients after injuries to peripheral nerves or facet-joint in rodents8,9,34,35,63. Similarly, several studies in neuropathic pain models demonstrated the ability of GBP to inhibit α2δ-1-induced26 or TSP-induced7,8,34,35 synaptogenesis. Most recently, GBP was also shown to inhibit TSP-2-induced synapse formation in purified culture of cortical neurons10. Despite the multidimensional evidence of GBP interference with TSP/α2δ-1 interaction, a direct GBP inhibition of this interaction on the molecular level has never been investigated before, to our knowledge. In the current study, we did not observe any inhibition of the direct TSP-4/α2δ-1SNTST interaction in the presence of increasing concentrations (up to 1 mM) of GBP (Fig. 4B). Furthermore, the highest GBP concentration used (1 mM) did not shift the TSP-4/α2δ-1SNTST binding curve (Fig. 4C). Although the utilised α2δ-1S NTST was mostly expressed as uncleaved form of the protein (in agreement with the original work describing a similar porcine α2δ-1 mutant40), we were able to demonstrate the ability of this α2δ-1S mutant to retain high affinity for GBP (Fig. 4A). For this purpose, a newly developed SPR-based binding assay suitable for detecting and quantifying the binding of small molecules to immobilized recombinant α2δ-1S was used. This SPR assay has the advantage of being radiolabel-free and can easily be used to determine the binding kinetics unlike the previously used 3H-GBP binding assay38,40,64,65. Taken together, our data confirmed that the proteolytic cleavage of α2δ-1 is not crucial for the formation of the GBP binding pocket40. The complete lack of GBP inhibition towards the interaction of purified TSP-4 with α2δ-1S NTST raises questions regarding the exact mechanism by which GBP can block the above-mentioned TSP-induced changes. It is possible that other unknown factors in the cellular environment are essential for GBP to interfere with α2δ-1/TSP-4 interaction and thereby mediating the known GBP inhibitory effects. Another possible explanation based on the recent findings of Chen et al.58,66 is that the α2δ-1/NMDA receptor complex, rather than α2δ-1/TSP-4 binding, represents the molecular target of gabapentinoid drugs to alleviate neuropathic pain.
Our efforts were as well focused on the investigation of the TSP-4/α2δ-1 interaction in a cellular system to get closer to the physiological/pathological situation in the CNS. We over-expressed full-length α2δ-1 in HEK293-EBNA cells and demonstrated both its intracellular and plasma membrane localisation in transfected cells (Fig. 5A). Treatment with increasing concentrations (up to 909 nM) of fluorescently labelled A555-TSP-4, however, showed no differences in binding of the protein to α2δ-1 overexpressing cells when compared to control cells (Fig. 5C, Supplementary Fig. S5). Furthermore, fluorescent signals of A555-TSP-4 did not co-localise with those of immunostained α2δ-1 on the cells (Supplementary Fig. S5). The observed loss of binding cannot be attributed to an impairment of the interaction of the two proteins by the fluorescent label of TSP-4 since the fluorescent protein was generated with a minimal dye-to-protein molar ratio and showing substantial α2δ-1S NTST binding in the ELISA-style assay (Fig. 5B). A possible explanation could instead be arising from the weak interaction of the two proteins under the conditions of the cell-based assay, unlike the ELISA-style assay. This means that very high local concentrations of TSP-4 in the proximity of cell-surface α2δ-1 would be required to enable the detection of their interaction by simulating the pathological situation (e.g. dramatic up-regulation following nerve injury). This could not, however, be achieved with the range of A555-TSP-4 concentrations (up to 909 nM) used in the assay. In our experiments, we over-expressed α2δ-1 in HEK293-EBNA cells without co-expression of α1 subunit, which interacts intracellularly with α2δ-1 before trafficking of the complex to the cell surface67. However, we assume that the lack of α1 subunit did not hamper the putative binding of TSP-4 to α2δ-1 on the cell surface. This assumption is based on recent data showing the ability of wild-type α2δ-1, expressed in HEK293 cells without co-expression of α1 subunits, to be efficiently transported to the cell surface and thereby become accessible to extracellular ligands like TSP10. Functionally, this α2δ-1 over-expression construct alone was able to rescue synapses in cortical organotypic slices from α2δ-1 knockout mice. This effect is found to be mediated through activation of the small Rho GTPase Ras-related C3 botulinum toxin substrate 1 (Rac1) and is independent of α1 subunits of the postsynaptic L-type calcium channels CaV1.2 and CaV 1.310.
The α2δ subunits are thought to promote membrane trafficking of the pore subunits of voltage-gated calcium channels17 and α2δ-1-driven allodynia in mice can be reversed by blockers of voltage-gated calcium channels like ω-conotoxin GVIA68. However, other findings suggest that the maladaptive changes contributing to chronic pain in rodents following nerve injuries and resulting from the interaction of dysregulated TSP-4 with α2δ-1 are partially independent of the role of the latter protein in regulating voltage-gated calcium channels’ trafficking and function50.
As previously mentioned, our data align with those of Lana et al.38 who reported no interaction of secreted TSP-4 with membrane-localised α2δ-1 on tsA-201 cells when subjected to immunocytochemical analysis. On the other hand, the same study showed weak intracellular interaction of both proteins in co-immunoprecipitation experiments from cells over-expressing both proteins38. It has therefore been postulated that the weak TSP-4/α2δ-1 interaction may occur in an intracellular compartment rather than on the cell surface38,69. This postulation is in contrast to the previous data showing synaptogenic effect of secreted TSPs which is mediated by neuronal α2δ-1 thought to be located either pre-8 or post-7,10synaptically. To our knowledge, there are no data so far showing the co-localisation of TSP(-4) and α2δ-1 in neuronal cell cultures or spinal cord tissue where TSP-induced changes (e.g. synaptogenesis) were demonstrated. To reveal the exact cellular localisation of TSP-4/α2δ-1 interaction it would be very helpful to simultaneously analyse co-immunostained TSP-4 and α2δ-1 proteins in cultures of neurons utilized in in vitro synapse assays6,7,10.
In summary, our results provide substantial in vitro biochemical evidence for a direct and specific Ca2+-insensitive TSP-4/α2δ-1 interaction which is rather weak. Importantly, GBP does not inhibit this interaction on a molecular level, indicating the possible involvement of other unknown factors or targets in mediating GBP inhibitory effects in neuropathic pain.
We, therefore, need to understand the exact and complete molecular mechanism of the TSP/α2δ‐1 interaction to really be able to design appropriate small molecule modulators – rather than being left to use and optimize the enigmatic properties of the serendipitously discovered gabapentinoid action.