
Calmodulin Supports TRPA1 Channel Association with Opioid Receptors and Glutamate NMDA Receptors in the Nervous Tissue

By Elsa Cortés-Montero, María Rodríguez-Muñoz, M. Carmen Ruiz-Cantero, Enrique J. Cobos, Pilar Sánchez-Blázquez, and Javier Garzón-Niño
Excerpt from the article published in International Journal of Molecular Sciences 22, no. 1: 229. Published: 28 December 2020, DOI: https://doi.org/10.3390/ijms22010229
Editor’s Highlights
- TRPA1 is the only mammalian member of the transient receptor potential (TRP) ankyrin channel subfamily.
- There is widespread agreement that TRPA1 plays an important role in chemonociception.
- mu-opioid receptors (MOR), via histidine triad nucleotide-binding protein 1 (HINT1) and type 1 sigma receptor (σ1R), connects with the NR1 subunit of NMDARs and positively regulates NMDAR calcium (Ca2+) fluxes.
- The HINT1 protein and calmodulin (CaM) bind to the N- and C-terminal domains of TRPA1.
- σ1R competes with CaM for binding to TRPs, except for its binding to the TRPA1 C-terminus, where σ1R promotes the binding of CaM.
- TRPA1 does not couple to MORs or NMDARs via HINT1 or the σ1R, thus the HINT1 protein or σ1R did not promote such associations but in the presence of CaM, the Nt and Ct cytosolic regions of TRPA1 coupled with the MOR and the NR1 subunit.
- The interaction of the TRPA1 channel with MORs and NMDARs relies on CaM.
- Mechanical allodynia caused by chronic sciatic nerve constriction injury (CCI) reduced the association of TRPA1 with MOR, DOR, and NMDARs.
- The association of TRPA1 channels with MOR, DOR, and NMDARs in different areas of the nervous system, as well as the plasticity of such associations, is in response to drugs and inflammatory and neuropathic pain.
Abstract
Transient receptor potential ankyrin member 1 (TRPA1) belongs to the family of thermo TRP cation channels that detect harmful temperatures, acids and numerous chemical pollutants. TRPA1 is expressed in nervous tissue, where it participates in the genesis of nociceptive signals in response to noxious stimuli and mediates mechanical hyperalgesia and allodynia associated with different neuropathies. The glutamate N-methyl-d-aspartate receptor (NMDAR), which plays a relevant role in allodynia to mechanical stimuli, is connected via histidine triad nucleotide-binding protein 1 (HINT1) and type 1 sigma receptor (σ1R) to mu-opioid receptors (MORs), which mediate the most potent pain relief. Notably, neuropathic pain causes a reduction in MOR antinociceptive efficacy, which can be reversed by blocking spinal NMDARs and TRPA1 channels. Thus, we studied whether TRPA1 channels form complexes with MORs and NMDARs that may be implicated in the aforementioned nociceptive signals. Our data suggest that TRPA1 channels functionally associate with MORs, delta opioid receptors and NMDARs in the dorsal root ganglia, the spinal cord and brain areas. These associations were altered in response to pharmacological interventions and the induction of inflammatory and also neuropathic pain. The MOR-TRPA1 and NMDAR-TRPA1 associations do not require HINT1 or σ1R but appear to be mediated by calcium-activated calmodulin. Thus, TRPA1 channels may associate with NMDARs to promote ascending acute and chronic pain signals and to control MOR antinociception.
1. Introduction
TRPA1 is the only mammalian member of the transient receptor potential (TRP) ankyrin channel subfamily [1,2]. Four identical subunits assemble to form the functional TRPA1 nonselective cation channel permeable to Ca2+, Na+and K+ [3]. The monomer contains six membrane-spanning domains and a presumed pore-forming region between the fifth and sixth transmembrane domains. The long N-terminal segment, which contains at least 16 ankyrin repeat domains as the shorter C-terminal region are predicted to be in the cytoplasm [4,5]. TRPA1 was initially suggested to function as a thermodetector of noxious cold stimuli [6]. Although this hypothesis remains controversial [7], there is widespread agreement that TRPA1 plays an important role in chemonociception. TRPA1 detects exogenous irritant compounds (e.g., formalin, acrolein, tear gas, chlorine, isothiocyanates and thiosulfinates from mustard, cinnamon, wasabi and garlic) and endogenous compounds (e.g., reactive oxygen species, bradykinin and 4-hydroxynonenal) produced during tissue damage or neurogenic inflammation [8,9,10,11]. Additionally, TRPA1 is activated by non-electrophilic molecules such as menthol and cannabinoids [12,13].
Painful peripheral stimuli are initially detected by primary afferent sensory nerve fibers (C and Aδ fibers) that have their soma in the trigeminal ganglia and the dorsal root ganglia (DRG). These peripheral neurons transmit this information to neurons within the spinal cord and thence to the brain via ascending neural circuits [8]. One of the most important sensory transducers that detects and transmits noxious stimuli is the transient receptor potential (TRP) ion channel family [11]. TRPA1 is expressed by nociceptive neurons of DRG and trigeminal ganglia, as well as in cutaneous primary afferent terminals [14]. Trpa1 mRNA has been localized to small- and medium-diameter sensory neurons, many of which are C- and Aδ-fiber nociceptors [12,15,16]. In the spinal dorsal horn, TRPA1 ion channels on the central terminals of peptidergic primary afferent nerve fibers regulate transmission to glutamatergic and GABAergic interneurons [17,18]. TRPA1 has been presented as a candidate to mediate inflammatory mechanical hyperalgesia as well as cold hyperalgesia under inflammatory conditions; indeed HC-030031, a TRPA1 selective antagonist, attenuates inflammatory- and neuropathy-induced mechanical hypersensitivity [19,20]. TRPA1 channels are also involved in the maintenance of mechanical and cold hyperalgesia in persistent inflammation [20]. Furthermore, formalin excites sensory neurons by directly activating TRPA1. In fact, blockade of TRPA1 in vivo, either using a specific antagonist or through disruption of the TRPA1 gene substantially attenuates pain-related responses to formalin [21]. Thus, based on its localization and functional properties, TRPA1 is considered a key player in acute and chronic (neuropathic) pain [22,23,24,25].
On the other hand, a series of neural changes associated with neuropathic pain originate from anomalous persistent activation of N-methyl-d-aspartate glutamatergic receptors (NMDARs), which triggers a cascade of events leading to central nervous system hypersensitization (“central windup”) [26,27]. In fact, neuropathic pain characterized by tactile allodynia and hyperalgesia remits after blockade of NMDAR function [28]. NMDARs in the spinal dorsal horn play a key role in chronic pain and central sensitization [29] and their interaction with opioid receptors plays an important role in regulating nociceptive transmission between primary afferents and dorsal horn neurons. NMDARs are present in primary afferents [30,31] and evoke the release of substance P from their central terminals [32]. Conversely, substance P release from primary afferents is inhibited by mu-opioid receptors (MORs) [33,34] and delta-opioid receptors (DORs) [35]. In the latent sensitization model of chronic pain, spinal cord hyperalgesia is maintained by NMDARs [36] and suppressed by MORs, DORs and kappa-opioid receptors [37]. In nervous tissue, NMDARs play an essential role in MOR desensitization [38,39] and neuropathic pain leads to reduced morphine efficacy and more rapid development of tolerance [40,41]. Interestingly, antinociceptive morphine tolerance is attenuated by blocking spinal TRPA1. This suggests that spinal TRPA1 may also contribute, at least in part, to the facilitation of morphine antinociceptive tolerance [42].
MOR, via histidine triad nucleotide-binding protein 1 (HINT1) and type 1 sigma receptor (σ1R), connects with the NR1 subunit of NMDARs [43,44] and positively regulates NMDAR calcium (Ca2+) fluxes [45]. σ1R binds to the NMDAR NR1 subunit in a Ca2+-dependent manner [44,46] and thus prevents the binding of the NMDAR negative regulator calmodulin (CaM) [47,48,49]. Notably, the HINT1 protein and CaM bind to the N- and C-terminal domains of various TRPs, including TRPA1. σ1R competes with CaM for binding to TRPs, except for its binding to the TRPA1 C-terminus, where σ1R promotes binding of CaM [50].
The aforementioned evidence prompted us to investigate whether TRPA1 channels associate with MORs, DORs and NMDARs. The study was performed principally on DRG, spinal cord and cortex and the possibility of direct associations was explored in in vitro assays with cloned sequences of these proteins. Ex vivo studies indicated the association of TRPA1 with MORs, DORs and NMDARs in vivo and these associations were altered by pharmacological interventions, the administration of a chemical irritant such as formalin and during neuropathic pain. The in vitro assays suggest a role for CaM in the association of TRPA1 with MORs and the NR1 regulatory subunit of NMDARs.
2. Results
2.1. TRPA1 Channel Association with MORs, DORs and NR1 Subunits, in the Cerebral Cortex, Spinal Cord and DRGs. Effect of Pharmacological Interventions
We confirmed the presence of TRPA1 channels in DRG, spinal cord and cerebral cortex of murine nervous tissue [4,16,25,51,52,53]. The 1125 residue monomer was immunodetected in synaptosome-enriched fractions using affinity-purified antibodies against the N-terminal region and an extracellular sequence. The opioid receptors MOR and DOR are present in the cortex and dorsal horn (substantia gelatinosa) of the spinal cord; they are also expressed on cell bodies of the sensory neurons (DRG) and are transported to their central terminals in the superficial dorsal horn and to peripheral terminals in peripheral tissues [54,55]. We observed that immunoprecipitated MOR and DOR were accompanied by TRPA1-related signals (Figure 1B), suggesting that they form functional complexes in nervous tissue. Similarly, the NR1 subunit of NMDARs coprecipitated TRPA1 in the spinal cord. The immunosignals mostly appeared as a doublet at approximately 140 and 150 kDa (Figure 1A), which is probably due to the described pair of N-glycosylation sites in the external loop of human TRPA1, which is flanked by the transmembrane regions TM1 and TM2 (721–960) [56]. Computer analysis indicated this possibility for murine TRPA1 [57] and in vitro, PGNase F abolished the 150 kDa band augmenting the intensity of the expected 140 kDa (Figure 1B).
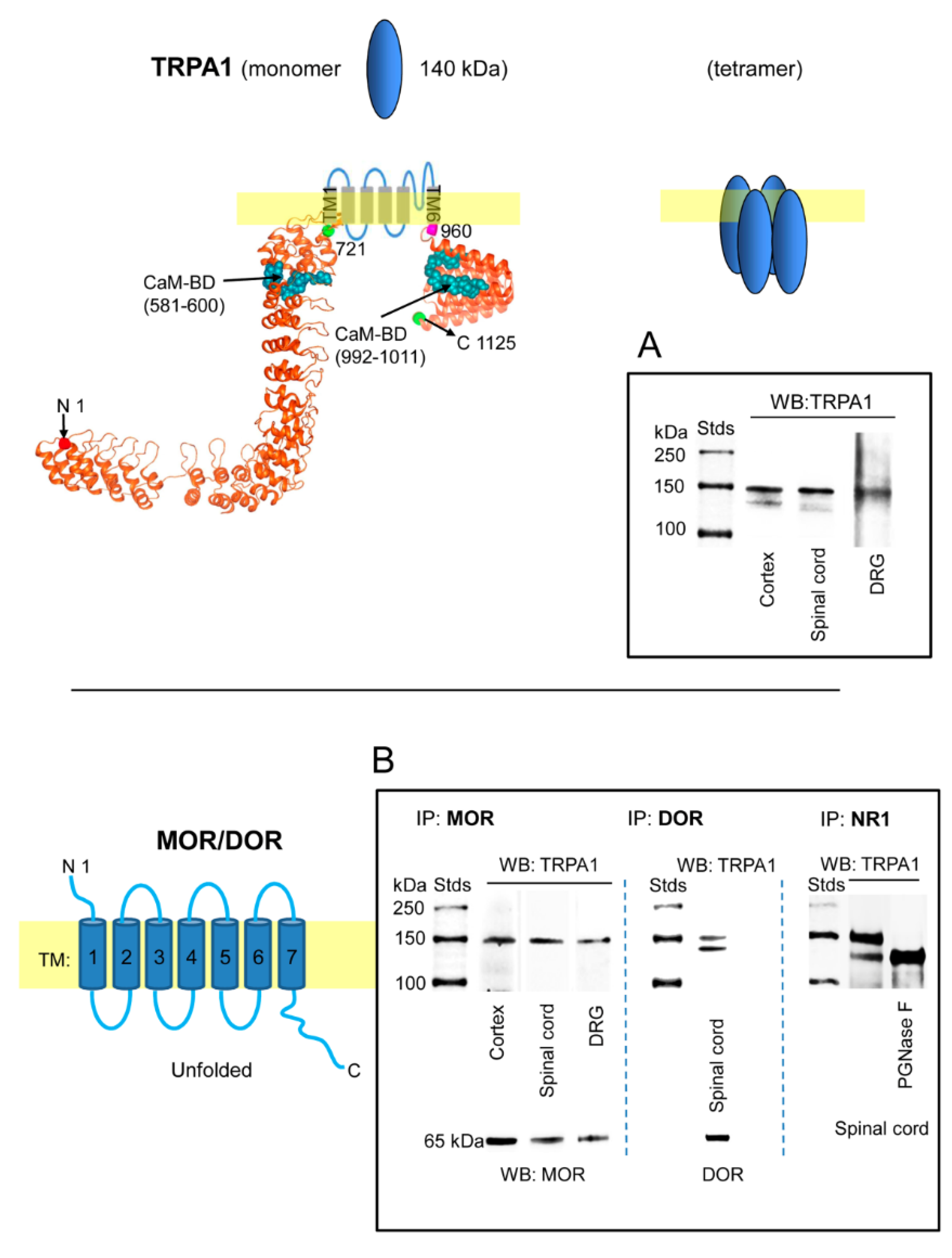
Expression of transient receptor potential ankyrin member 1 (TRPA1) channels in the cerebral cortex, spinal cord and dorsal root ganglia (DRG). TRPA1 association with mu-opioid receptors (MORs), delta-opioid receptors (DORs) and NR1 subunits. The structural model of the TRPA1 monomer was predicted by Novafold (DNASTAR Inc., Madison, WI, USA). The N- and C-terminal cytosolic sequences are linked by six transmembrane domains (TM). Ribbon model: The 3D structure of N- and C-terminal sequences shows the CaM-binding domains (BD) indicated by blue spheres. (A) Immunodetection of the TRPA1 monomer in synaptosomal fractions of murine cerebral cortex, spinal cord and dorsal root ganglia (DRG). (B) The membrane preparations were solubilized and the MOR, DOR and NR1 subunits of glutamate NMDARs were immunoprecipitated. The coprecipitation of the TRPA1 monomer was addressed by Western blotting. The expected size of the TRPA1 monomer is approximately 140 kDa; however, the immunosignals usually form a doublet of 140–150 kDa. The material associated with NR1 subunits was subjected to deglycosylation with PGNase F, which abolished the TRPA1 150 kDa band in favor of the 140 kDa band. Further details in Section 4.
The plasticity of protein complexes in response to pharmacological interventions and/or physiological alterations usually indicates their functional relevance. This possibility was addressed by injecting morphine, MOR agonist, via intracerebroventricular (icv; first or second ventricles) and intraperitoneal (ip) routes and NMDA, an activator of glutamate NMDARs, via the icv route. Morphine, at the dosage used, injected via the icv or ip route promoted similar levels of antinociception, approximately 75% of the maximum measurable effect in the analgesic test. The effect of the opioid peaked sharply at 30 min after icv injection, while antinociception was more sustained, with a maximum between 30 and 60 min, after ip administration. On the other hand, icv NMDA diminished the antinociception produced by supraspinal icv morphine (Figure 2A). We observed that icv-injection of NMDA increased MOR-TRPA1 and NR1-TRPA1 associations at the cortical level and the spinal cord level. On the other hand, icv morphine increased ex vivo coprecipitation of MOR-TRPA1 complexes at the cortex level and diminished that of NR1-TRPA1 at the two levels of the nervous system studied. Morphine injected via the ip route mostly modified both associations at the cortical level (Figure 2B).
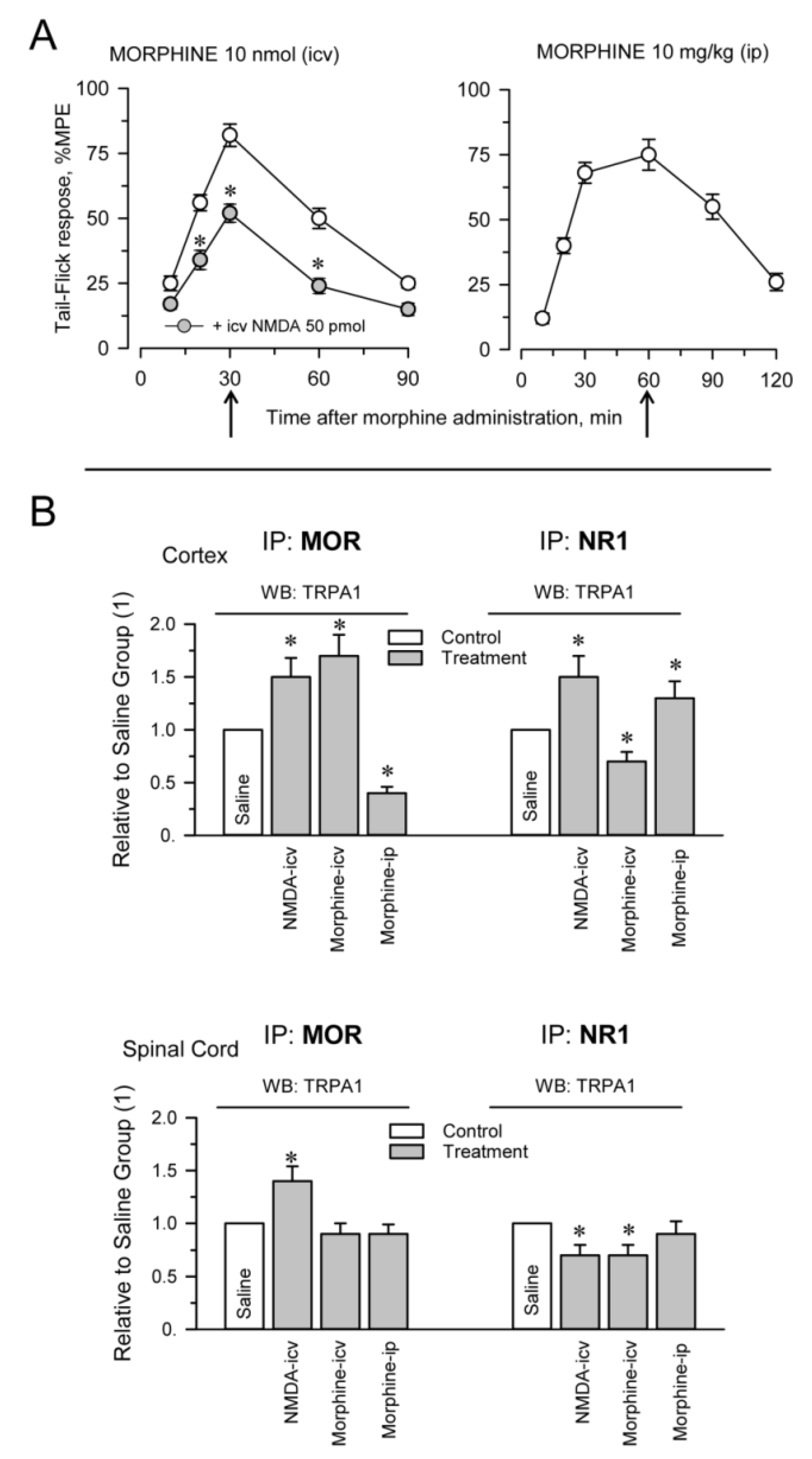
Pharmacological modulation of TRPA1 associations with MORs and glutamate NMDARs. Different groups of mice were icv-injected with saline, 50 pmol NMDA and 10 nmol morphine. Another group of mice received 10 mg/kg morphine via the ip route. (A) Antinociception promoted by morphine was determined in the warm water tail-flick test and is shown as a time-course. The effect of NMDA is shown relative to its antagonism of icv morphine analgesia. The points are the mean ± SEM from groups of six mice. * Significant differences with respect to the group that received only morphine icv, p < 0.05. (B) Effect of pharmacological interventions on the associations of TRPA1 with MORs and NR1 subunits. Thirty minutes after the icv injections and 60 min after ip morphine (indicated by the arrows), the mice were killed and cortical and spinal cord synaptosomal fractions were prepared. Following solubilization of these membrane preparations, MORs and NR1 subunits were immunoprecipitated. The coprecipitated TRPA1 was detached from the bait proteins and the presence of the monomer was analyzed by Western blotting. For each association and structure, namely, the cerebral cortex and spinal cord, the data were referred to the control group that received saline and was assigned an arbitrary value of 1. The bars are the mean ± SEM from two or three determinations. * Significant differences with respect to the control group, p < 0.05. Further details in Supplementary MaterialsFigure S4 and Section 4.
2.2. Inflammatory and Neuropathic Pain Alters TRPA1 Channel Association with MORs and NMDARs
The injection of formalin into the hind paw is widely used to evaluate the effects of analgesic compounds in laboratory animals. Intraplantar formalin excites sensory neurons by activating excitatory TRPA1 channels and this activity underlies the physiological and behavioral responses associated with this model of pain hypersensitivity, such as licking and biting responses [21,58]. The intraplantar injection of 0.5% formalin produced a biphasic behavioral reaction, with an initial phase lasting 5 min and a quiescent period, followed by a second less intense phase lasting from 10 to 25 min. Mice receiving saline, a solvent of formalin, did not show appreciable nociceptive-related responses during the time interval evaluated (Figure 3A). The mice did not exhibit pain-like behaviors toward the noninjected hind paw. We analyzed whether activation of TRPA1 channels on peripheral nerve terminals influences their association with MORs and NMDARs as well as the MOR-NR1 association in the spinal cord and DRG. Formalin almost completely abolished the MOR-NR1 association in DRG but increased this association in the spinal cord. The association of TRPA1 with MOR and NR1 subunits increased in the spinal cord and TRPA1-NR1 showed a tendency to augment in DRG (Figure 3B).
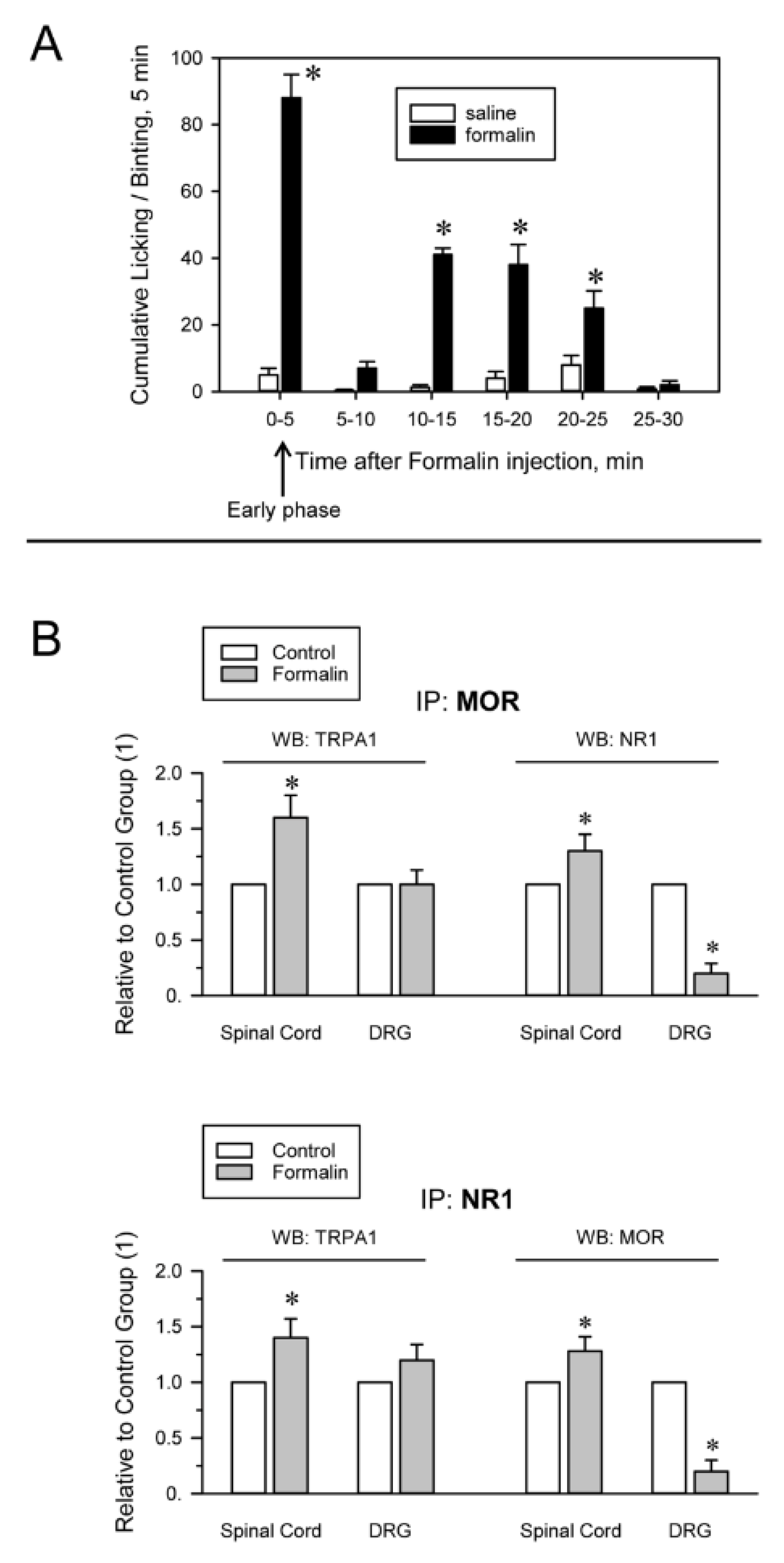
Formalin-induced inflammatory pain alters TRPA1 associations with MORs and NMDARs. (A) The mice received at time 0 an intraplantar injection of saline (control group) or 20 μL of a 0.5% formalin solution into one of the hind paws. The time spent licking/biting was recorded every 5 min by a blinded observer. The bars are the mean ± SEM from three determinations. * Indicates significant difference compared to the saline control group, p < 0.05. (B) The animals were sacrificed 5 min after the injection, when the early phase of formalin-induced pain had already peaked (indicated by the arrow). DRG and spinal cords were obtained. MORs and NR1 subunits were immunoprecipitated from the solubilized membrane preparations. At the end of the procedure, coprecipitated TRPA1 was detached from the bait proteins and analyzed by Western blotting * Indicates significant difference compared to the saline control group, p < 0.05. Further details in Figure 2, Supplementary Materials Figure S5 and Section 4.
The influence of neuropathic pain on the association of TRPA1 channels with opioid receptors and NMDARs was also evaluated. Neuropathic pain was induced by chronic sciatic nerve constriction injury (CCI). The nerve-injured mice maintained a healthy appearance and were well groomed and their body weight decreased after surgery but returned to preoperative values within 2–4 days. Seven days after surgery, CCI mice but not sham-operated mice showed allodynia and on days 12 to 15, the nociceptive responses of CCI mice nearly returned to pre-surgery levels [59] (Figure 4A). The Sham and CCI mice were sacrificed on day 7 and the nervous structures were obtained for immunoprecipitation studies. Since cortical NMDAR-MORs associations remain unchanged in the murine CCI model of neuropathic pain [41], we analyzed the extent of their interaction in the pons-medulla and periaqueductal gray matter (PAG). We observed reductions in the MOR-TRPA1 association of ≥50% in the pons-medulla, PAG and spinal cord. Similarly, the DOR-TRPA1 and NR1-TRPA1 associations were significantly diminished in the spinal cord of CCI mice (Figure 4B).
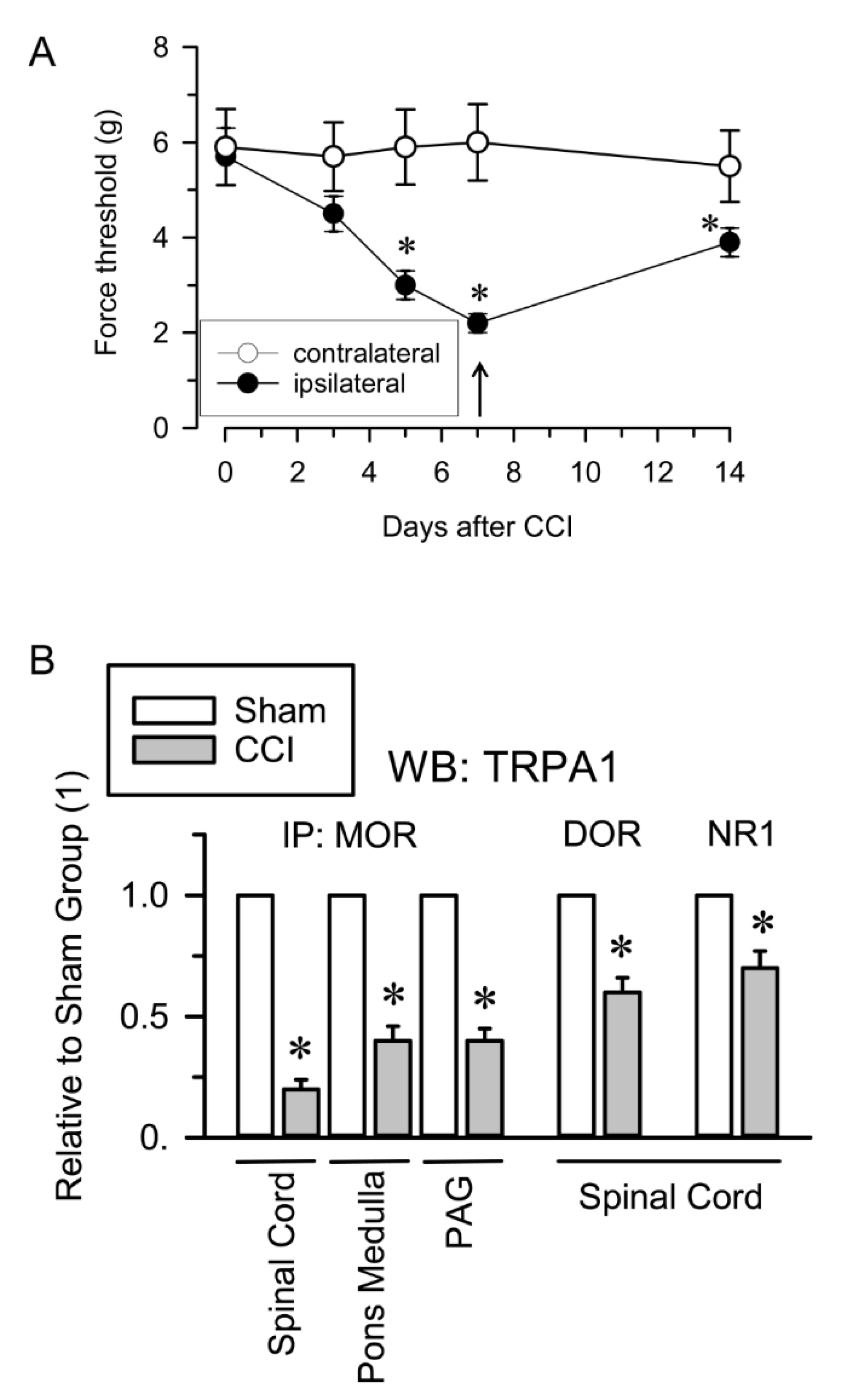
TRPA1 associations with opioid receptors and NMDARs in the CCI model of neuropathic pain. (A) Induction of mechanical allodynia. Chronic constriction injury (CCI) of the sciatic nerve caused neuropathic pain in mice. The paw withdrawal thresholds of the contralateral and ipsilateral paws of the mice were measured before (indicated as 0) and 3, 5, 7 and 14 days after surgery. The force (in grams) at which the mice withdrew their paws in response to von Frey hair stimulation was determined as an index of mechanical allodynia. The data are the mean ± SEM of six mice. * Indicates significant difference compared to the nociceptive threshold of the sham-operated control group on day 0 (7th after surgery); p < 0.05. (B) Seven days after surgery, sham and CCI mice were killed and synaptosomal fractions from different neural areas were obtained. The MOR, DOR and NR1 subunits were immunoprecipitated from the solubilized membrane preparations and coprecipitated TRPA1 was analyzed by Western blotting. * Indicates significant difference compared to the sham control group, p < 0.05. Further details in Figure 2, Supplementary Materials Figure S6 and Section 4.
Because NMDARs connect with MORs via HINT1 proteins and σ1Rs [46,60], we explored this possibility for the associations of TRPA1 channels with MORs and NMDARs. We evaluated this possibility in P2 fractions from the spinal cords of HINT1−/− and σ1R−/− mice. In these mice, the associations of TRPA1 with these proteins were comparable to those of the corresponding wild-type mice (Supplementary Materials Figure S1).
2.3. Calmodulin Mediates TRPA1 Channel Association with MORs and NR1 Subunits of Glutamate NMDARs
In in vitro assays, we observed that MOR does not bind to cytosolic TRPA1 Nt or Ct regions and that 3 mM CaCl2does not promote this association (Figure 5). In this paradigm, σ1R failed to couple TRPA1 Nt or Ct to the MOR and the HINT1 protein also failed to bride MOR to TRPA1 Nt (Supplementary Materials Figure S2). Because HINT1 does not bind to TRPA1 Ct, this assay was not extended to this region [50]. However, in the presence of Ca2+-activated CaM, TRPA1 Nt and Ct coupled to MORs (Figure 5). CaM binds to TRPA1 Ct in the absence of Ca2+ and this association increases at a physiological 3 mM concentration of Ca2+ [50]. Notably, the MOR-CaM interaction showed a similar pattern of dependence on Ca2+ and the MOR-TRPA1 Ct association was observed even in the absence of Ca2+ (Supplementary Materials Figure S3). The NR2A and NR2B subunits of NMDARs form stable associations with regulatory α2δ1 proteins [61], which may block the access of these subunits to TRPA1 channels. Thus, we evaluated the possibility that the cytosolic region of the NR1 subunit interacting with TRPA1 monomers. We observed that NR1 does not interact directly with TRPA1 Nt or Ct but in the presence of 3 mM Ca2+, CaM facilitated these interactions (Figure 6).
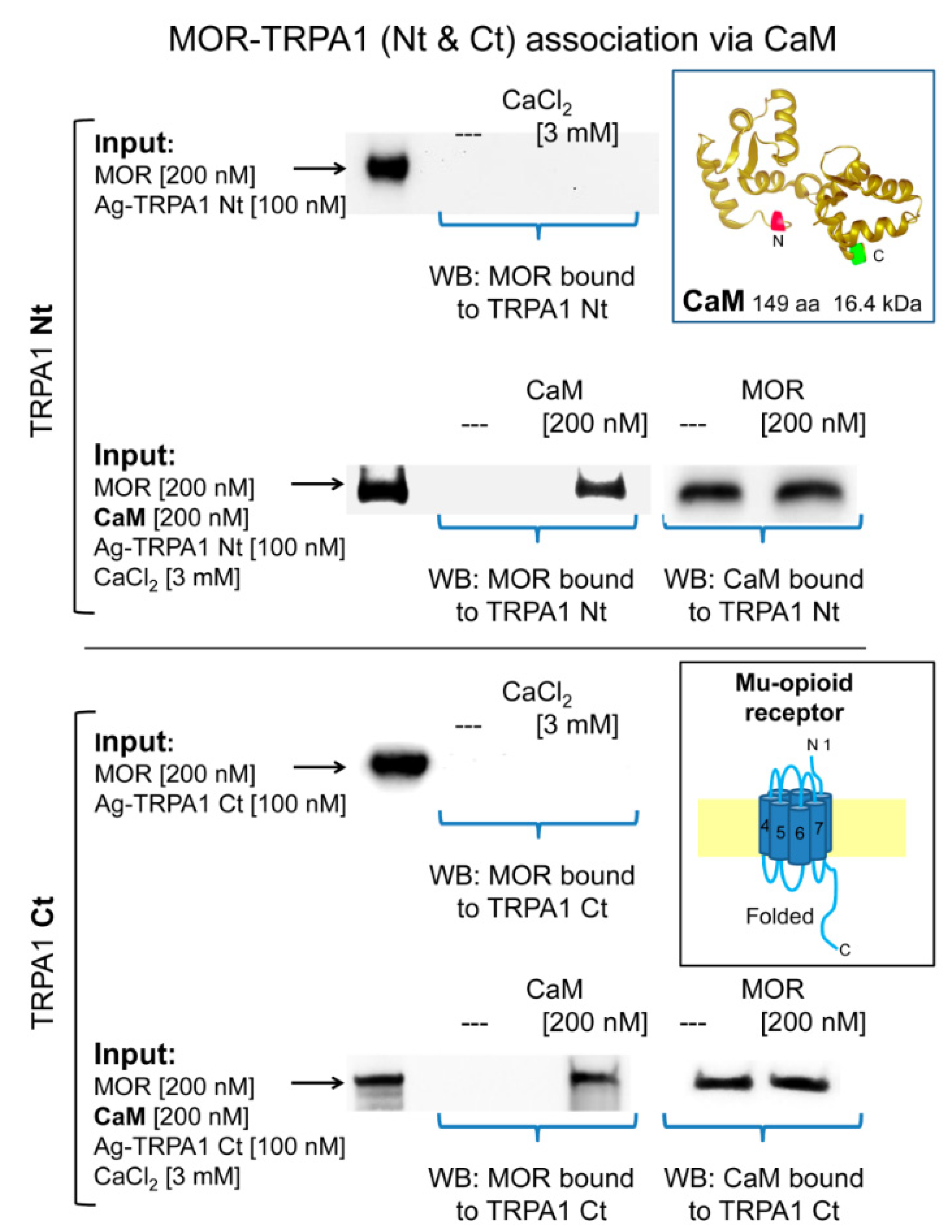
Calmodulin mediates the association of TRPA1 with MORs. The recombinant cytosolic Nt and Ct regions of TRPA1 were covalently attached to N-Hydroxysuccinimide (NHS)-agarose beads and then incubated with MOR in the absence and presence of 3 mM CaCl2. In another set of assays, the TRPA1 regions were sequentially incubated with calmodulin (CaM) and MORs in the presence of 3 mM CaCl2. At the end of the incubation, agarose-TRPA1 was recovered by several cycles of washing-resuspension and the bound proteins were detached with 2× Laemmli buffer and resolved by SDS-PAGE followed by immunodetection. Further details in the Methods. Key: Input, Proteins incubated with agarose TRPA1, MORs are immunodetected; WB, immunodetected protein in Western blot.
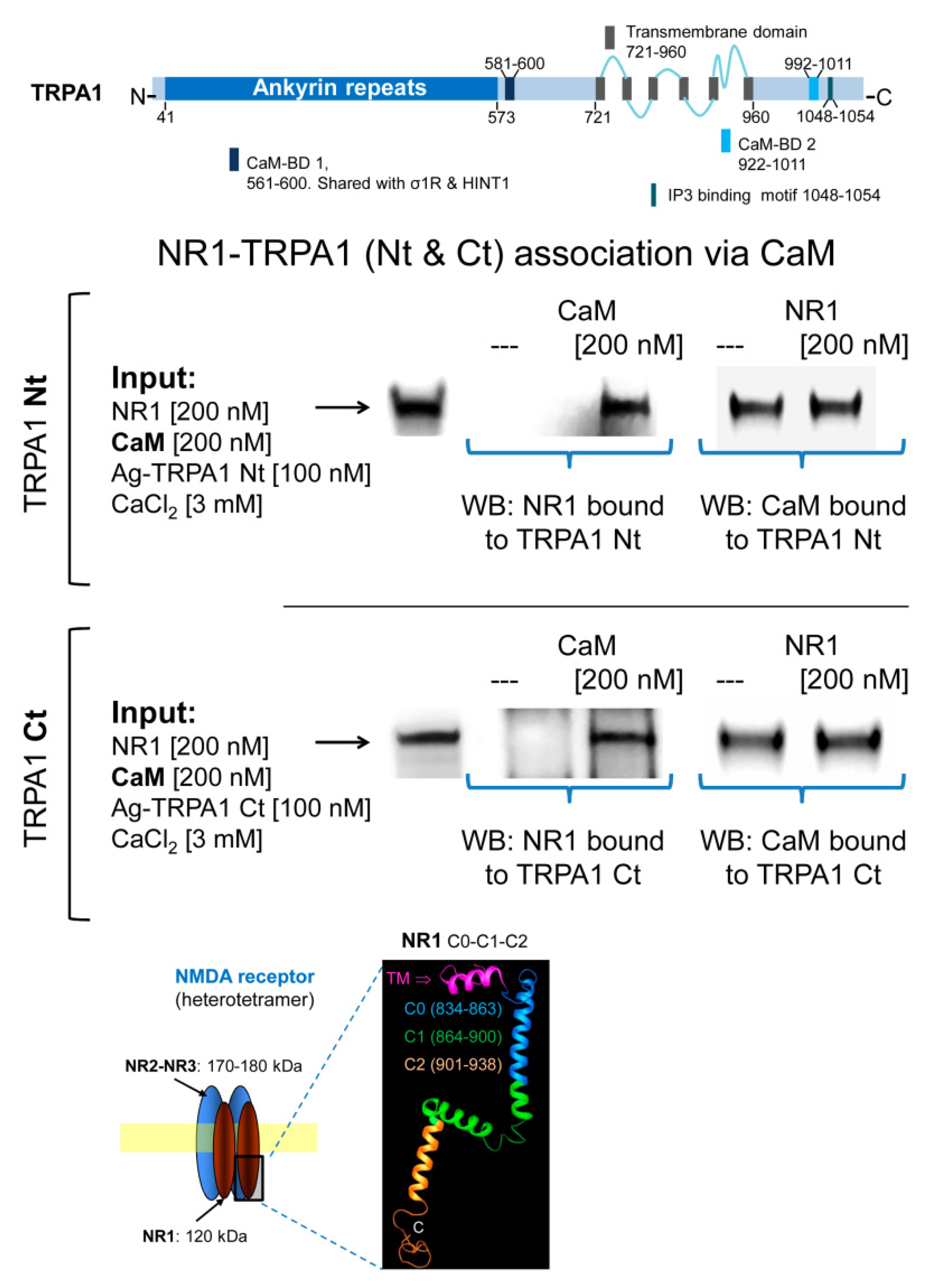
Calmodulin mediates the association of TRPA1 with NMDARs. Linear representation of TRPA1 monomer showing CaM- and IP3-binding domains (BD) [5]. Recombinant cytosolic Nt and Ct regions of TRPA1 were covalently attached to NHS-agarose beads and then were sequentially incubated with CaM and NR1 subunits of NMDARs in the presence of 3 mM CaCl2 and 30 µM of a peptide mapping the C1 region of NR1 subunit (879–888: TFRAITSTLA) [46]. CaM and NR1 associated with TRPA1 regions were detected by Western blotting. Details in Figure 5 and Section 4.
3. Discussion
This study suggests that a relationship exists between TRPA1 and glutamate NMDARs. Both cationic channels participate in the genesis of noxious signals, are functionally coupled to opioid receptors such as the MOR and form associations, which are modulated by their activity.
The TRPA1 channel was first cloned from human fetal lung fibroblasts and is also expressed in other types of nonneuronal cells, including keratinocytes, melanocytes, mast cells, odontoblasts, enterochromaffin cells and β-cells of the Langerhans islets [62,63,64]. In murine peripheral sensory neurons, TRPA1 is expressed in peptidergic (TRPV1+) neurons [4]. In the spinal dorsal horn, TRPA1 on the central terminals of peptidergic primary afferent nerve fibers regulates transmission to glutamatergic and GABAergic interneurons [12,15,16]. In the central nervous system, TRPA1 is also present in the somatosensory cortex, hippocampus, pons-medulla and spinal cord [52,53,65,66].
Our study identified the presence of TRPA1 channels in the synaptosomal fraction of murine frontal cortex, PAG, pons-medulla, spinal cord and DRG, where MORs and DORs are also expressed. The finding that TRPA1 and MOR/DOR coprecipitate suggests a functional relationship between the nociceptor channel and these antinociceptive G protein-coupled receptors (GPCRs). Glutamate NMDARs also associate with MORs via NR1 subunits of NMDARs that carry the regulatory cytosolic C1 segment [43]. This association is regulated by HINT1 proteins and σ1Rs, with the latter competing with the NMDAR inhibitor CaM [47] for binding to NR1 subunits [44,46]. The activation of MORs recruits the activity of NMDARs, which exerts negative feedback on MOR signaling. This mechanism mostly operates for opioids such as morphine, which promotes limited internalization of MORs and mediates the short-term tolerance that follows the effects of high doses of opioids [48].
In the resting state, the association of MORs with NMDARs elevates the threshold for direct NMDAR activation and facilitates the recruitment of this glutamatergic activity in response to MOR signaling [43,48,60]. The MOR-NMDAR association is supported by HINT1 and σ1Rs and accordingly, in mice lacking expression of these proteins, the MOR-NMDAR association greatly diminishes [44,67]. Our data indicate that TRPA1 does not couple to MORs or NMDARs via HINT1 or the σ1R and thus the TRPA1-MOR and TRPA1-NR1 associations were maintained in neural P2 fractions obtained from HINT1−/− and σ1R−/− mice. In in vitro assays, the HINT1 protein or σ1R did not promote such associations but in the presence of CaM, the Nt and Ct cytosolic regions of TRPA1 coupled with the MOR and the NR1 subunit. The TRPA1 monomer binds HINT1, σ1R and CaM via the N-terminal region and σ1R and CaM but not HINT1 via the C-terminal region. Calcium promotes CaM and σ1R binding to TRPA1 Nt and Ct but CaM also binds to TRPA1 Ct in the absence of Ca2+. Moreover, σ1R competes with CaM at the TRPA1 Nt but promotes CaM binding to the Ct region [50]. The regulation of TRPA1 activity by CaM appears to be complex and thus, CaM mediates Ca2+-dependent potentiation and inactivation of the channel. TRPA1 contains a noncanonical CaM binding domain (CaMBD) in the C-terminal region, while the N-terminal region, which exhibits weak CaM binding, is critical for Ca2+-induced desensitization [68,69]. The CaMBD in the N-terminus is close to the linker domain and/or pre-S1 helix, regions structurally near the C-terminal CaMBD [5], which is consistent with previous studies demonstrating that extracellular Ca2+ regulates TRPA1 by acting through a channel site that needs to be very close to the channel pore [16]. Thus, the topology of the CaMBDs on TRPA1 makes the simultaneous interaction of CaM with the TRPA1 channel and cytosolic regions of the MOR and NR1 subunits of NMDARs feasible.
At low activation levels, MORs couple with NMDARs and TRPA1 channels and the activation of MORs promoted by icv morphine diminishes the MOR-NMDAR association [46] but enhances the MOR-TRPA1 association at the supraspinal level. Under low Ca2+ influx, TRPA1 Ct and MORs bind to CaM and thus, these signaling proteins may form basal complexes. The activation of TRPA1 channels, for example, by formalin, may augment CaM binding; hence, the MOR-TRPA1 association will increase, as observed for the spinal cord. When the MOR is activated, the interplay between σ1R and CaM at the TRPA1 Nt will diminish CaM binding and thus the MOR association but at the TRPA1 Ct, σ1R always promotes CaM binding, which supports MOR association. After icv morphine, the latter situation was observed at the cortical level but not at the spinal level, probably because icv-injected morphine barely reaches spinal MORs. On the other hand, ip-injected morphine produced the opposite effect, diminishing the MOR-TRPA1 association at the cortex but not at spinal level. Morphine, when injected by the systemic route, crosses the blood brain barrier but mostly acts at the peripheral and spinal levels [70]. The opioid receptors, MOR and DOR, are present in the dorsal horn (substantia gelatinosa) of the spinal cord, cell bodies of sensory neurons (DRG), central terminals in the superficial dorsal horn and peripheral terminals in peripheral tissues [54,55]. Although, ip morphine did not alter TRPA1-MOR association in the spinal cord, the reduction in these complexes observed in the cortex may originate at the MORs located in sensory neurons reaching the upper areas through the neuromodulation of ascending pathways.
MOR-activated NMDARs participate in a negative feedback loop that dampens MOR signaling; however, there is no evidence that activated TRPA1 channels promote such an effect. On the other hand, GPCRs such as type 1 mGluR5 increase TRPA1 sensitivity and this regulation requires the activity of PKA and PKC on specific residues of the channel monomer [71]. Similarly, in DRG, bradykinin B2 and the PAR2 receptor can also stimulate TRPA1 function, probably via Gq/G11-PLC-PKC [51,72]. The TRPA1 residues identified as targets for PKA, PKC and Cdk5 phosphorylation are outside the CaMBDs [71,73,74] and thus should not interfere with CaM binding or CaM-mediated associations of the channel with additional partner proteins. In this scenario, MORs, by inhibiting sensory neuron excitability [75], may oppose TRPA1 channels; however, MOR activity may also contribute to TRPA1 activity via PKC, PKA and Cdk5 [48]. Indeed, the MOR can contribute zinc ions and nitric oxide (NO) to the regulation of TRPA1 channels. The C-terminal region of the MOR couples via HINT1 proteins to regulators of GTPase signaling (RGS) proteins 20 and 17 (RGSZ1 and RGSZ2), which carry zinc ions bound to cysteine rich domains (CRDs). In the MOR-mediated complex, the N-terminal region of neural nitric oxide synthase (nNOS) binds to these RGSZ proteins. Upon activation of MORs, nNOS generates NO and releases zinc ions from RGSZ CRDs [48,49]. Because, intracellular zinc ions activate TRPA1 through interactions with Cys641 and Cys1021 in the C-terminal fragment [76], MORs may increase TRPA1 activity, providing zinc ions but it may also reduce its activity by removing Cys-bound zinc ions via NO.
NMDARs are present on nociceptive primary afferents from the skin [77,78,79] and trigeminal ganglia [80]. Approximately 47% of unmyelinated peripheral axons are immunopositive for NMDARs and are believed to be involved in synaptic transmission between primary afferents and dorsal horn neurons [81,82]. NMDARs play a relevant role in central sensitization in spinal neurons and are also involved in the release of specific neurotransmitters during pain transmission at both the peripheral and central terminals of sensory neurons [83]. Thus, NMDA receptors may be involved in functionally divergent nociceptive systems [84]. The activation of MORs or the direct activation of NMDARs by icv NMDA brings about reductions in the MOR-NR1 association [43]. These situations recruit protein kinases such as PKC, PKA and CaMKII [48], which, via the phosphorylation of MOR cytosolic residues, exert negative feedback on opioid signaling and disrupt the MOR association with NMDARs. The binding of Ca2+-CaM to NR1 reduces the probability of NMDAR channel opening, while phosphorylation of the NR1 cytosolic region by PKC and PKA blocks access to CaM, increases the binding of σ1R and promotes NMDAR function [46,47].
In contrast, the interaction of the TRPA1 channel with MORs and NMDARs relies on CaM. Supraspinal administration of NMDA increased the MOR-TRPA1 association at the cortical and spinal levels and promoted or diminished NMDAR-TRPA1 associations at the cortical and spinal level, respectively. NMDA-activated NMDARs increase cytosolic Ca2+ and thus, may augment the binding of supraspinal TRPA1 channels with the aforementioned proteins via Ca2+-activated CaM. This mechanism may trigger the changes observed at the spinal level via modulation of descending pathways, which finally affect TRPA1 association with MORs and NMDARs in the opposite direction. Formalin also brought about increases in TRPA1 association with MORs and NMDARs, particularly in the spinal cord and caused a great reduction in the MOR-NMDAR association in DRG, with values comparable to those produced by CCI in the spinal cord, pons-medulla and PAG [41]. These changes may be mediated by the activity of NMDARs and indeed, NMDAR antagonists attenuate formalin-induced pain behaviors [84], suggesting that the glutamatergic receptor is necessary for TRPA1 nociceptive signals to propagate. The icv or ip activation of MORs also affected the association between TRPA1 and NMDARs, probably by recruiting the activity of NMDARs [43,46]. Morphine icv, reduced cortical and spinal TRPA1-NMDAR associations and ip morphine increased this association at the cortical level.
The associations of TRPA1 with these signaling proteins show a more uniform change profile as a consequence of neuropathic pain. Mechanical allodynia caused by CCI reduced the association of TRPA1 with MOR, DOR and NMDARs. Disruption of the MOR-NMDAR association is also observed in this animal model of neuropathy [41]. Thus, adaptive responses to the sustained overactivity of NMDARs “winding up” differ from those underlying acute pharmacological interventions and inflammatory formalin-induced pain. In summary, our study has shown the association of TRPA1 channels with MOR, DOR and NMDARs in different areas of the nervous system, as well as the plasticity of such associations in response to drugs and inflammatory and neuropathic pain. These findings may provide new avenues for understanding TRPA1 physiology in relation to signaling proteins relevant to pain perception.