
Alterations in Excitatory and Inhibitory Synaptic Development Within the Mesolimbic Dopamine Pathway in a Mouse Model of Prenatal Drug Exposure
By Taylor Boggess, James C. Williamson, Ethan B. Niebergall, Hannah Sexton, Anna Mazur, Richard D. Egleton, Lawrence M. Grover, and W. Christopher Risher
Excerpt from the article published in Frontiers in Pediatrics , 9, 13 December 2021, DOI: https://doi.org/10.3389/fped.2021.794544
Editor’s Highlights
- Neonatal abstinence syndrome (NAS) is a collection of signs and symptoms commonly observed in the newborns of mothers who abused certain types of drugs during their pregnancy.
- Prenatal drug exposure alter reward circuitry and addiction-like behavior in adolescents and adults.
- Excitatory and inhibitory synaptic populations are significantly disrupted by prenatal exposure to either the opioid buprenorphine or gabapentin, separately or in combination, and that these disruptions are strongly influenced by α2δ-1 function.
- Haploinsufficiency of the calcium channel subunit α2δ-1, a critical player in developmental synapse formation and maturation, would result in further disruption of these synaptic populations.
- α2δ-1 is the binding site for a class of astrocyte-secreted extracellular matrix glycoproteins known as thrombospondins (TSPs), which promote synapse formation and maturation during early development.
- Significant drug treatment effects on synaptic development are shown with the loss of just a single copy of α2δ-1, which may indicate that astrocyte-mediated synaptogenic pathways were disrupted with the drug treatment paradigms.
- Changes in synapse number may indicate a general increase in net excitation in response to rewarding stimuli. α2δ-1, is involved in critical synaptogenic pathways mediated by astrocytes, was also shown to have an important role in the changes in synapse formation induced by prenatal drug exposure.
Abstract
The rise in rates of opioid abuse in recent years in the United States has led to a dramatic increase in the incidence of neonatal abstinence syndrome (NAS). Despite improved understanding of NAS and its acute symptoms, there remains a paucity of information regarding the long-term effects of prenatal exposure to drugs of abuse on neurological development. The primary goal of this study was to investigate the effects of prenatal drug exposure on synaptic connectivity within brain regions associated with the mesolimbic dopamine pathway, the primary reward pathway associated with drug abuse and addiction, in a mouse model. Our secondary goal was to examine the role of the Ca+2channel subunit α2δ-1, known to be involved in key developmental synaptogenic pathways, in mediating these effects. Pregnant mouse dams were treated orally with either the opioid drug buprenorphine (commonly used in medication-assisted treatment for substance use patients), gabapentin (neuropathic pain drug that binds to α2δ-1 and has been increasingly co-abused with opioids), a combination of both drugs, or vehicle daily from gestational day 6 until postnatal day 11. Confocal fluorescence immunohistochemistry (IHC) imaging of the brains of the resulting wild-type (WT) pups at postnatal day 21 revealed a number of significant alterations in excitatory and inhibitory synaptic populations within the anterior cingulate cortex (ACC), nucleus accumbens (NAC), and medial prefrontal cortex (PFC), particularly in the buprenorphine or combinatorial buprenorphine/gabapentin groups. Furthermore, we observed several drug- and region-specific differences in synaptic connectivity between WT and α2δ-1 haploinsufficient mice, indicating that critical α2δ-1-associated synaptogenic pathways are disrupted with early life drug exposure.
Introduction
Neonatal abstinence syndrome (NAS) is a collection of signs and symptoms commonly observed in the newborns of mothers who abused certain types of drugs during their pregnancy. The developing fetus acquires a physiological dependence on these drugs and, after being separated from the supply of drug at birth, the infant soon displays the symptoms of withdrawal, which can include irritability, tremors, excessive crying, poor feeding, diarrhea, and, in some of the more severe cases, seizures (1). The rise in the incidence rate of NAS in recent years (2, 3) corresponds with the rise in rates of opioid abuse among pregnant women (4, 5) and the United States in general (6). Considerable research efforts have been made to increase the understanding of the pathophysiology of NAS as well as methods to improve treatment of NAS symptoms (1, 7–10). However, knowledge concerning the long-term effects of prenatal exposure to drugs of abuse on neurological development is limited (11).
Opioids remain one of the most widespread and commonly abused classes of drug among the mothers of NAS patients. One of the most commonly used opioid drugs among pregnant substance abuse patients is buprenorphine, a partial μ-opioid receptor agonist that is often prescribed to replace and prevent the reinforcing effects of more harmful and addictive opioid drugs, such as heroin or fentanyl. Pregnant mothers who are addicted to opioids are commonly enrolled in medication-assisted treatment (MAT), which combines less-addictive prescription opioid drugs—namely methadone or buprenorphine—with behavioral counseling in order to treat addiction and achieve better health outcomes for the mother and child. However, emerging evidence asserts that any opioid use during pregnancy, even when part of a medical treatment plan, can have deleterious effects on the developing fetus (12, 13).
While the increased prevalence of opioid abuse is often credited with the rise in incidence of NAS, it is important to note that many mothers of NAS patients abuse multiple drugs besides, or in combination with, opioids. One prescription drug that has been shown to be increasingly co-abused along with opioids is gabapentin, an anticonvulsant drug also prescribed for the treatment of neuropathic pain (14). Gabapentin was initially believed to have no potential for abuse or addiction (14, 15). However, surveys of opioid use disorder patients have found that as many as 26% of those interviewed reported abusing gabapentin for nonmedical reasons, often abusing gabapentin in combination with opioids as a means to potentiate the experienced high (14, 16, 17). In addition, clinicians have observed a unique presentation of NAS in infants whose mothers abused both opioids and gabapentin while pregnant, with symptoms including tongue thrusting, back arching, and increased eye wandering (18).
Although the mechanisms by which gabapentin and opioids may interact with each other are unknown, the addictive nature of such drugs of abuse can largely be attributed to their activity within the mesolimbic dopamine pathway (Figure 1) (19–21). Also known as the dopamine reward pathway, this pathway functions by encoding certain experiences (e.g., food, sex, drugs) as pleasurable, then reinforcing future behaviors in an attempt to seek out those same rewarding experiences. Opioids are known to interact with this reward pathway by binding to opioid receptors on the surfaces of interneurons within the ventral tegmental area (VTA), a central location for the cell bodies of a large number of dopaminergic neurons. This opioid-receptor interaction inhibits the release of GABA within the VTA, thus disinhibiting dopamine release by dopaminergic neurons (22). The axons of VTA dopaminergic neurons project to the nucleus accumbens (NAC), where increased phasic release of dopamine in response to external stimuli has been shown to reinforce pleasurable behaviors including eating food, engaging in sexual activity, or taking drugs of abuse (23). The anterior cingulate cortex (ACC) is involved in a variety of functions, including reward anticipation and modulation of goal-oriented motor activity (24, 25) and has been shown to influence motivation via interaction with the mesolimbic dopamine pathway (26), with projections from the VTA having been shown to release dopamine within the ACC in response to microinjections of a μ-opioid receptor agonist (27). The prefrontal cortex (PFC) is highly associated with decision making and impulse control and has been shown to have axonal projections to the NAC capable of modulating neuronal activity within this region (28). Projections from the medial PFC to the NAC have been shown to be involved in reinstatement of drug-seeking behavior in response to stress or drug-associated cues (29, 30).
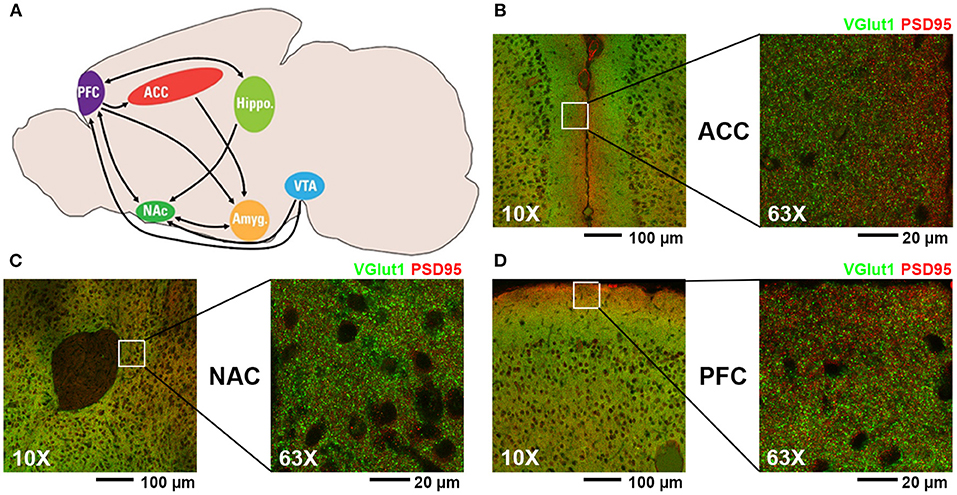
(A) Diagram of mouse brain regions associated with mesolimbic dopamine pathway (PFC, prefrontal cortex; ACC, anterior cingulate cortex; NAC, nucleus accumbens; VTA, ventral tegmental area; Hippo, hippocampus; Amyg, amygdala). (B–D) Representative fluorescent confocal microscopic images of excitatory synaptic staining (presynaptic VGlut1, green; postsynaptic PSD95, red) at 10X and 63X magnification of the regions analyzed in this study: (B) ACC, (C) NAC, and (D) PFC.
Examining the effects of drugs of abuse (specifically opioids and gabapentin) on synaptogenesis within brain regions associated with the mesolimbic dopamine pathway may lead to insight on how these drugs affect neurological development in prenatally exposed children. In this study, we used fluorescence immunohistochemistry (IHC) in a mouse model of early life drug exposure to investigate the effects of both buprenorphine and gabapentin on developmental synaptic connectivity. Our hypothesis was that exposure to buprenorphine, gabapentin, or a combination of both drugs during critical periods for synaptic development would lead to significant disruptions in excitatory and inhibitory synaptic connectivity in brain regions associated with addiction and reward. We further hypothesized that haploinsufficiency of the calcium channel subunit α2δ-1, the receptor for gabapentin and a critical player in developmental synapse formation and maturation, would result in further disruption of these synaptic populations.
…
Results
Confirming Drug Accumulation in Neonatal Mouse Brain Tissue
Each day during the dosing period, pregnant mice were administered drug treatments as 1 ml of a 1:1 sweetened condensed milk/water solution served in a plastic dish. Dishes from the previous day were always observed to be empty, confirming that the dams routinely consumed the entirety of the treatment solution. To confirm drug passage from the dam to the pups, the brains of newborn pups from litters prenatally treated with buprenorphine or gabapentin were isolated, lysed, and subjected to liquid chromatography-tandem mass spectrometry to determine the brain tissue concentrations of each drug. Both buprenorphine and gabapentin were detected in the brains of the prenatally exposed pups (Table 1), prompting further investigation of brain connectivity changes as a result of the presence of these compounds.
Body Mass of Dams and Pups
In order to produce the required number of WT and α2δ-1 heterozygous (+/–) pups for IHC in this study, the following litter numbers were required: 4 litters dosed with vehicle control, 5 litters dosed with 30mg/kg gabapentin, 4 litters dosed with 5mg/kg buprenorphine, and 5 litters dosed with both drugs. Immediately before anesthetization and perfusion at P21, dams and all pups from each litter were weighed and their weights were recorded. The mean masses for the dams of the different litters and the pups in each sex-genotype-treatment group were calculated (Table 2). The mean body masses for dams revealed a significant main effect of treatment, F(3, 14) = 50.21, p < 0.0001. Dams treated with buprenorphine (p = 0.0122), gabapentin (p = 0.0006), and buprenorphine + gabapentin (p < 0.0001) were all significantly lower than the mean body mass of the vehicle control dams. Significant differences were also observed among the various pup groupings across treatment [F(3, 42) = 140.4, p < 0.0001], genotype [F(1, 42) = 12.66, p =0.0009], treatment × sex [F(3, 42) = 4.487, p = 0.0081], and genotype × sex [F(1, 42) = 5.627, p = 0.0223]. No significance was found for the main effect of sex [F(1, 42) = 0.5268, p = 0.4720] or the interactions of treatment × genotype [F(3, 42) = 1.301, p = 0.2866] or treatment × genotype × sex [F(3, 42) = 2.039, p = 0.1229]. Full multiple comparisons are listed in Supplementary Table 1. Of note, key differences between treatments that reached significance included: WT male vehicle vs. either WT male buprenorphine (Bup) or WT male buprenorphine plus gabapentin (BupGBP), WT female vehicle vs. WT female BupGBP; heterozygous (Het) male vehicle vs. either Het male Bup or Het male BupGBP, and Het female vehicle vs. Het female BupGBP.
Early Life Exposure to Buprenorphine Increases Excitatory Synapses in the Mesolimbic Dopamine Pathway
We next wanted to determine whether excitatory synaptic development within the mesolimbic dopamine pathway was significantly impacted by the presence of buprenorphine and/or gabapentin in utero. Fluorescence IHC followed by confocal imaging was used to distinguish and quantify excitatory glutamatergic synapses within three different brain regions of WT mouse pups at P21 (Figure 2). Synapses were identified by the co-localization of presynaptic VGlut1 and postsynaptic PSD95 (Figure 2A). This type of imaging-based synapse analysis relies on the fact that these markers are expressed in completely different neuronal compartments (i.e., VGlut1 in axons, PSD95 in dendrites), and would only appear co-localized in confocal microscopy when directly opposed at sites of synaptic contact. Indeed, rotating the channels out of alignment with each other results in a dramatic decrease in co-localized puncta (Supplementary Figure 1). Beginning with excitatory glutamatergic synapses within the ACC (Figure 2B), we observed a significant main effect between groups, H = 99.00, p < 0.0001 (full list of multiple comparisons reported in Supplementary Table 3), as mice treated with either buprenorphine or buprenorphine in combination with gabapentin had a significantly increased mean number of synapses than mice treated with vehicle control. Interestingly, gabapentin treatment on its own did not significantly impact excitatory synapse number compared to vehicle, despite its known role as an inhibitor of prominent synaptogenic pathways (34). Similar results were also observed within the NAC (Figure 2C; H = 119.2, p < 0.0001), with buprenorphine and buprenorphine together with gabapentin having significantly more synapses than vehicle, with no significant effect of gabapentin alone. However, in the NAC, the combination drug treatment group also exhibited a significantly higher number of synapses than the gabapentin-only group. In contrast, within the PFC (Figure 2D; H = 165.0, p < 0.0001), no significant differences were observed among any of the treatment groups when compared to vehicle.
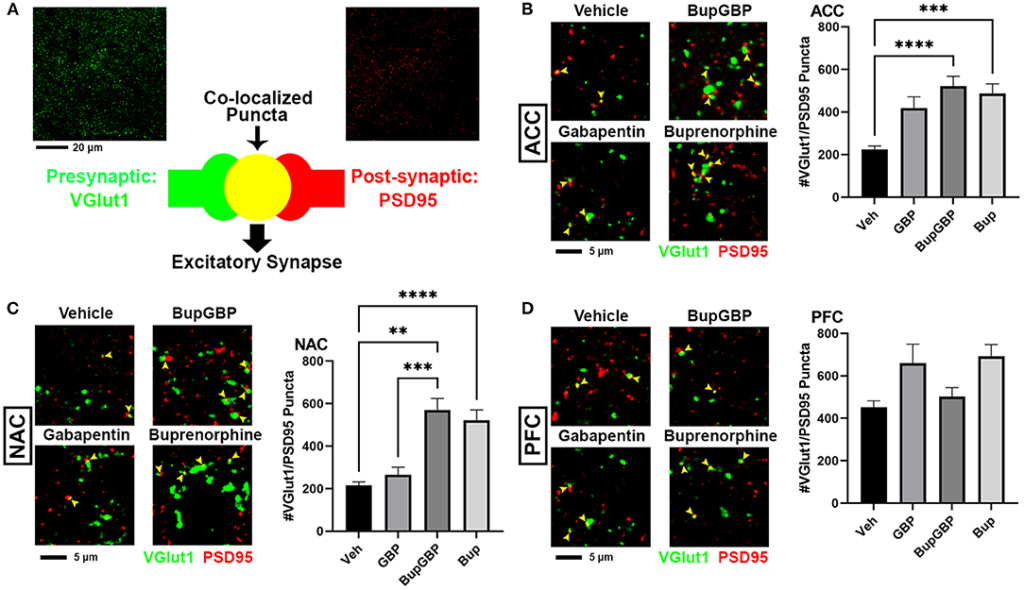
Increased excitatory synapses with early life buprenorphine exposure. (A) Diagram illustrating co-localization of puncta representing presynaptic (VGlut1, green) and postsynaptic (PSD95, red) fluorescent antibody label pairs for quantifying excitatory glutamatergic synapses. (B–D) Representative IHC images (left) and quantification (right) of co-localized VGlut1 (green) and PSD95 (red) excitatory synaptic puncta (yellow arrowheads) from prenatal drug-exposed WT C57Bl/6J P21 mouse brain within (B) the anterior cingulate cortex (ACC), (C) nucleus accumbens (NAC), and (D) prefrontal cortex (PFC). n = 7 (vehicle control), 6 (GBP), 7 (Bup+GBP), 6 (Bup); **p < 0.01; ***p < 0.001; ****p < 0.0001.
Combined Buprenorphine/Gabapentin Exposure Decreases Inhibitory Synapse Number
The same IHC procedure used for glutamatergic excitatory synapses was next used to stain for GABAergic inhibitory synapses (identified by the co-localization of presynaptic VGAT and postsynaptic gephyrin; Figure 3A) within the same brain regions of the previously analyzed WT pups. As with excitatory synapses, we observed a main effect of treatment, H = 196.5, p < 0.0001 (full list of multiple comparisons reported in Supplementary Table 4), for inhibitory synapses within the ACC (Figure 3B). A significant decrease in mean number of inhibitory synapses was observed in the combination drug treated mice compared to all other treatment groups. As with excitatory synapses, similar observations were made in the NAC (Figure 3C; H = 219.8, p < 0.0001), with the buprenorphine plus gabapentin group having significantly fewer inhibitory synapses than the other groups. The PFC, however, was the only region that exhibited an increase in inhibitory synapse number with any of the drug treatments (Figure 3D; H = 121.8, p < 0.0001), with the buprenorphine treatment group having significantly more VGAT/gephyrin co-localized puncta than the vehicle control group. Yet, as in the ACC and NAC, the combination treated mice had significantly fewer synapses within the PFC than mice from either of the single drug treatment groups, but not compared to vehicle control.
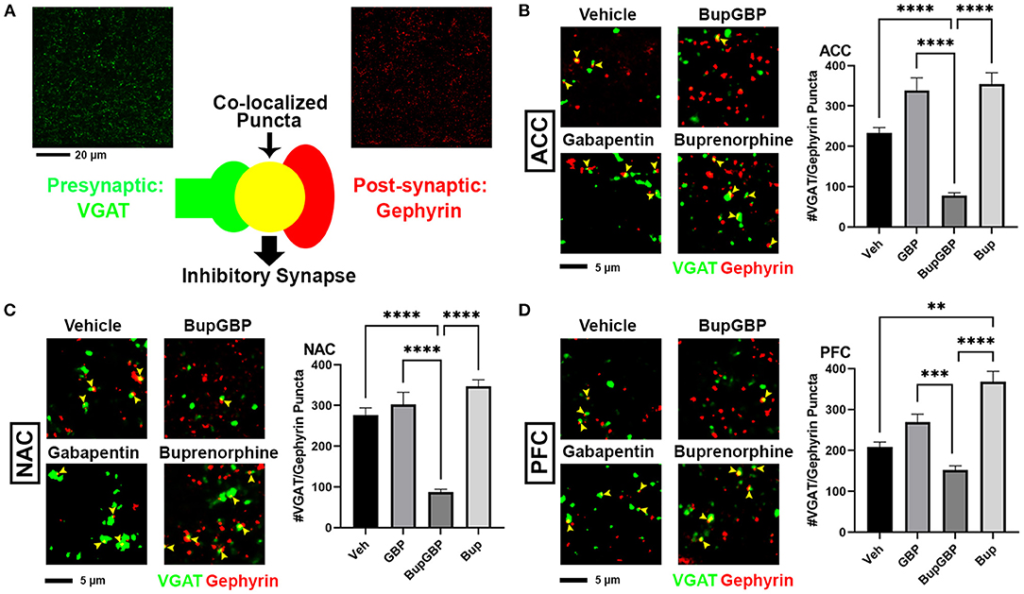
Dual exposure to buprenorphine and gabapentin decreased inhibitory synapses at P21. (A)Diagram illustrating co-localization of puncta representing presynaptic (VGAT, green) and postsynaptic (PSD95) fluorescent antibody label pairs for quantifying inhibitory GABAergic synapses. (B–D)Representative IHC images (left) and quantification (right) of co-localized VGAT (green) and gephyrin (red) inhibitory synaptic puncta (yellow arrowheads) from prenatal drug-exposed WT C57Bl/6J P21 mouse brain within (B) ACC, (C) NAC, and (D) PFC. n = 7 (vehicle control), 6 (GBP), 7 (Bup+GBP), 6 (Bup); **p < 0.01; ***p < 0.001; ****p < 0.0001.
α2δ-1 Haploinsufficiency Results in Region- and Treatment-Specific Differences in Synaptic Connectivity
Gabapentin has previously been shown to inhibit excitatory synapse formation by binding to the neuronal calcium channel subunit α2δ-1 (34), and this mechanism is proposed to underlie the efficacy of gabapentin in the alleviation of symptoms in both epilepsy and neuropathic pain (35, 36). We previously showed deficits in synaptic connectivity in the brains of young mice lacking α2δ-1 (31). To determine whether α2δ-1-mediated synaptic development is impacted by early life drug exposure, we expanded our synapse analysis to compare our WT synapse numbers with those from mice that were haploinsufficient for α2δ-1. Comparing excitatory synaptic development, within the ACC (Figure 4A; H = 99.00, p < 0.0001), only the gabapentin treated α2δ-1 +/- mice showed a significantly greater number of synapses compared with WT mice that received the same treatment. A similar gabapentin-induced increase was observed within the NAC (Figure 4B; H = 119.2, p < 0.0001). The same held true for the α2δ-1 +/- PFC (Figure 4C; H = 165.0, p < 0.0001), which also exhibited more synapses than WT mice with buprenorphine treatment.
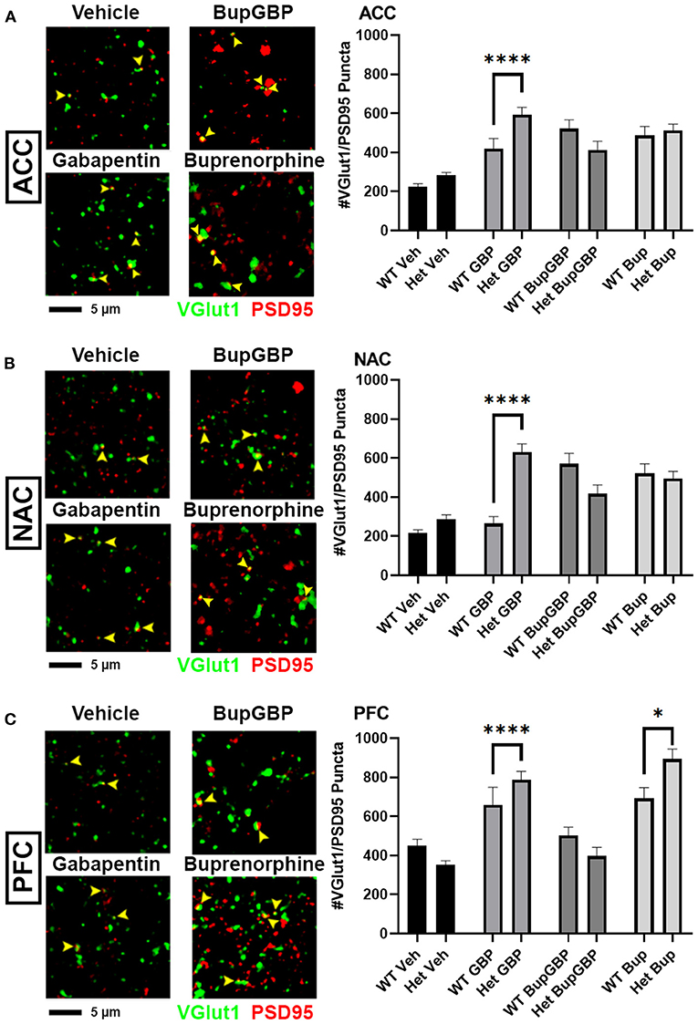
Altered excitatory synaptic connectivity following prenatal drug treatment in α2δ-1 haploinsufficient mice. Representative IHC images (left) of co-localized VGlut1 (green) and PSD95 (red) excitatory synaptic puncta (yellow arrowheads) from prenatal drug-exposed α2δ-1 +/- (Het) C57Bl/6J P21 mouse brain with quantification (right) compared to WT C57Bl/6J P21 mouse brains with the same prenatal treatment within (A) ACC, (B) NAC, and (C) PFC. Het: n = 11 (vehicle control), 7 (GBP), 7 (Bup+GBP), 7 (Bup); *p < 0.05; ****p < 0.0001.
Compared to excitatory synaptic development, much less is known about the roles of α2δ-1 and gabapentin in the formation of inhibitory circuits. We concluded our analyses by comparing mean inhibitory synapse numbers in α2δ-1 +/- mice to WT mice across the 4 different treatment groups (Figure 5). In the vehicle control, gabapentin, and buprenorphine groups, α2δ-1 +/- mice had significantly fewer synapses within the ACC compared to their WT counterparts (Figure 5A; H = 196.5, p < 0.0001). Only in the combination treatment group did we observe a significantly higher number of synapses in the ACC of α2δ-1 +/- mice compared to WT. Within the NAC (Figure 5B; H = 219.8, p < 0.0001), a similar trend was observed among the vehicle control, gabapentin, and buprenorphine groups as was seen in the ACC, with significantly fewer VGAT/gephyrin co-localized synapses in α2δ-1 +/- mice than WT. However, in the NAC, the α2δ-1 +/- mice in the combination treatment group did not differ significantly from the WT mice. Finally, within the PFC (Figure 5C; H = 121.8, p < 0.0001), the most dramatic significant difference was observed in the buprenorphine treatment group, with α2δ-1 +/- mice showing significantly fewer inhibitory synapses than WT mice. Taken together, these results indicate that α2δ-1 plays an important regulatory role in both excitatory and inhibitory synaptic development in the context of early life drug exposure.
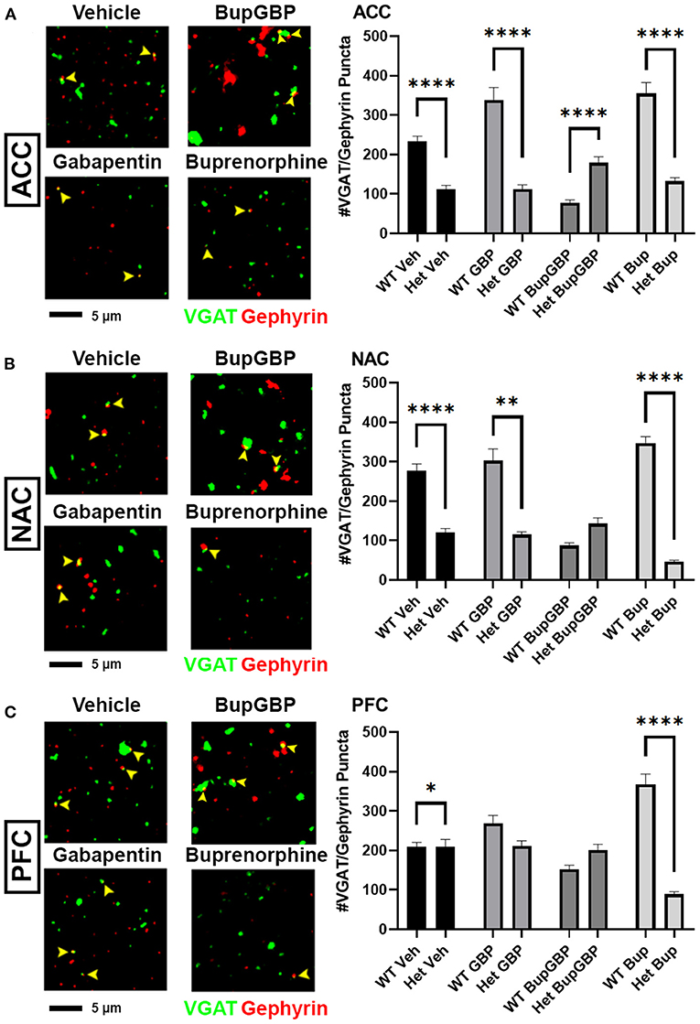
α2δ-1-influenced changes in inhibitory synapse development in drug-exposed pups. Representative IHC images (left) of co-localized VGAT (green) and gephyrin (red) inhibitory synaptic puncta (yellow arrowheads) from prenatal drug-exposed α2δ-1 Het C57Bl/6J P21 mouse brain and quantification (right) compared to WT C57Bl/6J P21 mouse brains with the same prenatal treatment within (A) ACC, (B)NAC, and (C) PFC. Het: n = 11 (vehicle control), 7 (GBP), 7 (Bup+GBP), 7 (Bup); *p < 0.05; **p < 0.01; ****p< 0.0001.
Discussion
Glutamatergic signaling has been shown to be critical to many different aspects of opioid abuse and addiction (37). Previous studies have shown that morphine-induced activation of dopaminergic neurons cannot occur within the VTA without glutamatergic modulation (38), and glutamate release within the NAC has been shown to be associated with heroin addiction and reinstatement of heroin-seeking behavior in rats (39). In addition, GABAergic synaptic activity has also been heavily implicated in the acute response to drugs and the reinforcement of drug seeking behavior (40, 41). However, relatively little is known about how exposure to drugs of abuse during critical developmental periods affects the trajectories of these synaptic populations. The goal of this study was to investigate the effects of early life drug exposure on synaptic development within brain regions associated with addiction and reward, as well as to explore the role of the gabapentin receptor, α2δ-1, in these alterations of synaptogenesis. Our results indicate that excitatory and inhibitory synaptic populations are significantly disrupted by prenatal exposure to either the opioid buprenorphine or gabapentin, separately or in combination, and that these disruptions are strongly influenced by α2δ-1 function.
Significant increases in excitatory glutamatergic synapses were observed in multiple regions associated with the mesolimbic dopamine reward pathway in WT mice that had undergone prenatal exposure to only buprenorphine or buprenorphine in combination with gabapentin. Concomitantly, mice exposed to the combination buprenorphine-gabapentin treatment in utero showed significant decreases in inhibitory GABAergic synapses compared to the vehicle control group within these same regions. In addition, the combined impact of buprenorphine and gabapentin resulted in significantly lower numbers of inhibitory synapses compared to either single drug treatment in all three brain regions, suggesting a synergistic effect of polysubstance use on synaptic development. Given the increased prevalence of polysubstance abuse in mothers of infants born with NAS, as well as the worsened clinical conditions observed in many of these children (42, 43), this finding is of particular concern. Taken together, these results indicate a net increase in excitatory glutamatergic signaling capability within the mesolimbic dopamine pathway of mice prenatally exposed to buprenorphine and gabapentin. Glutamatergic neurotransmission is heavily implicated in many aspects of drug use, including acute effects, consolidation, craving/seeking, withdrawal, and relapse [reviewed in Heinsbroek et al. (37)]. Whether the synaptic population shifts observed in this study result in greater dopamine release in response to rewarding stimuli, such as drugs of abuse, or a greater risk for developing substance abuse disorder later in life remains to be elucidated.
Drug Dosing Paradigm Correlates
The dosing for buprenorphine in this study was in line with a study by Martin et al. that showed 5 mg/kg to be capable of producing significant increases in pain threshold in rats (44). The gabapentin dosage was chosen based on the finding that 30 mg/kg gabapentin was sufficient to produce analgesia in mice (45). The dosing schedule, starting on E7 and ending on P11, was chosen to correspond to a period of significant synaptogenesis within mice (46–48), a period which also roughly correlates to the second trimester of human fetal development (49, 50). At birth, a mouse pup is considered to be at a developmental stage similar to that of a human fetus in the late second trimester. Despite the fact that newborn mouse pups are no longer receiving direct exposure to drugs of abuse via the placental blood supply, there were still precedents in the literature for postnatal drug transference to the brains of pups being nursed by drug-exposed dams. Kongstorp et al. demonstrated that buprenorphine accumulates and remains in the brain tissue of newborn rodents for several days following birth (51). In addition, both buprenorphine and gabapentin have been shown to pass from mother to infant via breast milk, albeit in low concentrations compared to the mothers’ blood plasma levels (52–54). We confirmed the presence of both buprenorphine and gabapentin in the brains of prenatal drug exposed pups, though the levels in our study were admittedly at the lower ends of the ranges reported in the referenced works. Regardless, the results of this study show that the chosen treatment paradigm was capable of disrupting normal CNS synaptic development in mouse pups. The fact that we detected relatively lower concentrations in our animals may even suggest that the observed deficits may have been further exacerbated if the accumulated drug levels had been higher.
Gabapentin, α2δ-1, and Astrocytes
Unexpectedly, single drug treatment with gabapentin did not result in any significant differences in excitatory or inhibitory synapses within any of the examined brain regions in the WT. This finding would appear to contradict the findings of previous studies showing that gabapentin inhibits normal excitatory synapse development by interfering with pathways mediated by α2δ-1. Though originally designed as a structural analog for the neurotransmitter GABA, gabapentin does not bind to GABA A or GABA B receptors. It is instead proposed that gabapentin decreases neurotransmitter release from the presynaptic terminal by inhibiting Ca+2 influx through L-type channels (55, 56). Gabapentin performs this action by binding to α2δ-1 on the surface of neurons at the site of synaptic terminals (35, 57). α2δ-1 is also the binding site for a class of astrocyte-secreted extracellular matrix glycoproteins known as thrombospondins (TSPs), which promote synapse formation and maturation during early development (31, 34). It may be possible that, despite interference by gabapentin in normal TSP/α2δ-1 mediated synaptic development, the net level of synaptogenesis is maintained via other signaling mechanisms as a means to compensate for the gabapentin-induced decrease in glutamate release.
In an attempt to elucidate the impact of α2δ-1 on synaptic development in the drug exposed brain, the original goal of this study was to mate α2δ-1 +/- males and females to produce pups of all three possible α2δ-1 genotypes, i.e. α2δ-1 +/- (Het), α2δ-1 +/+ (WT), and α2δ-1 -/- (knockout; KO). However, after several attempts to breed α2δ-1 KO pups, we found that such pups were either not being born or did not survive to age P21, particularly in the buprenorphine or gabapentin treatment groups. Given the widespread presence of α2δ-1 throughout the mouse CNS (58, 59), skeletal muscle (60, 61), and cardiac muscle (62), the global knockout of this Ca2+ channel subunit, in combination with exposure to drugs of abuse during fetal development, may have been potentially lethal.
Despite the difficulty in generating the necessary number of drug exposed α2δ-1 KO pups for this study, we still observed significant drug treatment effects on synaptic development with the loss of just a single copy of α2δ-1, which may indicate that astrocyte-mediated synaptogenic pathways were disrupted with the drug treatment paradigms. Indeed, it has been previously shown that opioids can directly dysregulate the expression and secretion of synaptogenic TSP by astrocytes (63, 64). Far from being a passive bystander in the CNS, astrocytes have been increasingly shown to influence and maintain the excitatory/inhibitory balance necessary for homeostasis (65). Not only are astrocytes able to mediate both excitatory (47) and inhibitory (66) synaptogenesis, they are also able to mediate synapse elimination via phagocytosis mediated by astrocytic cell surface receptors such as MEGF10 and MERTK (67). Outside of synapse regulation, astrocytes are also capable of both uptake (68–70) and release (71, 72) of glutamate and GABA. μ-opioid receptors have been confirmed on the surface of astrocytes within the hippocampus, NAC, and VTA (73), while morphine and other μ-opioid receptor agonists have been shown to alter DNA synthesis (74) and increase expression of glial fibrillary acidic protein (GFAP), an indicator of activation and reactivity, in astrocytes (75). Greater understanding of the role of astrocytes in maintaining excitatory/inhibitory synaptic balance, coupled with increased awareness of the effects of drugs of abuse on astrocytes during development, may facilitate novel insight into how neurological function may be impacted in cases of early life drug exposure.
Limitations and Conclusions
In this study, we used a mouse model of early life drug exposure to examine the effects such exposure may have on synaptic development within brain regions traditionally associated with reward and addiction. Though there were key regional differences, an overall increase in glutamatergic excitatory synapses as well as a general decrease in GABAergic inhibitory synapses was observed in the mesolimbic dopamine pathway of mice prenatally exposed to buprenorphine or buprenorphine in combination with gabapentin. Such changes in synapse number may indicate a general increase in net excitation in response to rewarding stimuli. α2δ-1, which binds to gabapentin and is involved in critical synaptogenic pathways mediated by astrocytes, was also shown to have an important role in the changes in synapse formation induced by prenatal drug exposure, as evidenced by the significant differences observed between WT and α2δ-1 haploinsufficient animals. We were not able to determine the effects of prenatal drug exposure on mice completely lacking expression of α2δ-1 (possibly due to increased lethality when combined with drug exposure), nor did our study investigate whether these structural synaptic findings persist to later ages and/or correlate with altered physiological function or behavior. However, this work may serve to inform future studies examining these functional effects as well as more longitudinal studies that may link history of prenatal drug exposure with altered reward circuitry and addiction-like behavior in adolescents and adults.