
Nociceptive TRP Channels and Sex Steroids

By Óscar Enciso-Pablo, Karina Angélica Méndez-Reséndiz, Tamara Rosenbaum, and Sara Luz Morales-Lázaro
Excerpt from the article published in Reproductive Hormones, edited by Courtney Marsh. London: IntechOpen, March 24th, 2021. DOI: 10.5772/intechopen.95552
Editor’s Highlights
- Several experiments made in pain animal models have demonstrated the existence of sexual dimorphism in response to pain. Rats from a diabetic neuropathy model exhibited lower mechanical pain thresholds than males.
- Androgens act as direct molecular regulators of some nociceptive ion channels function, playing dual roles as nociceptive and antinociceptive molecules.
- Progesterone impacts on nociception by regulating the expression of TRPV1 channels.
- This regulation is through a non-classical mechanism that requires an intermediate protein, the Sig-1R, which is a dynamic chaperone mainly localized in the endoplasmic reticulum and that can also be mobilized to the plasma membrane and nuclear envelope.
- Progesterone decreases the levels of channels localized in the cell surface in HEK293 cells by disrupting a protein complex formed between TRPV1 channels and the Sig-1R and by affecting the proper folding of TRPV1.
- Progesterone also disrupts the protein complex formed between Sig-1R and the TRPA1 and TRPM8 channels, suggesting that this steroid could impact the nociception through these nociceptive TRP channels.
- These data open a research field in which we will deepen our knowledge in the role of molecular sexual dimorphism and the interplay between nociceptive TRP channels and sex steroids.
Abstract
Proteins belonging to Transient Receptor Potential (TRP) family are nonselective cation channels that play an essential role in mammalian physiology, functioning as transducers of several environmental signals including those of chemical, thermal and mechanical natures. A subgroup of these receptors is expressed in sensory neurons where they are activated by noxious stimuli and are key players of pain responses in the organism. Some TRP channels are molecular targets for the classical and non-classical effects of sex steroids. This chapter will describe the close relationship between nociceptive TRP channels and sex steroids as well as their impact on nociception and pain-related responses.
1. Introduction
The somatosensory system is a complex network of neurons and peripheral receptors that encodes specific information about the state of organisms and their environment, providing them with the ability to detect harmful stimuli [1, 2, 3].
According to the International Association for the Study of Pain (IASP), the neuronal process encoding noxious stimuli is known as nociception. A subpopulation of high-threshold sensory neurons known as nociceptors mediates this process by detecting harmful signals from chemical and physical nature Figure 1 [1, 4]. Nociceptors are neurons with a peripheral axonal branch that innervates cutaneous, articular and visceral afferents, and a central axonal branch that innervates the dorsal horn of the spinal cord. The cell bodies of mammalian nociceptors are located in the dorsal root ganglia (DRG) and in the trigeminal ganglia (TG) and they are classified as medium and small diameter neurons. The axons of these nociceptors are classified as Aδ- and C-fibers depending on their diameter, degree of myelinization, conduction speed and the type of sensory stimuli that they transmit [1, 4, 5].
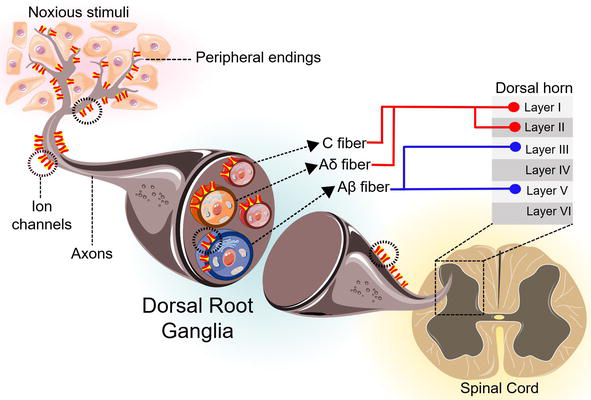
Nociception. Harmful stimuli are transduced along the terminal axon of specialized pseudounipolar neurons (nociceptors), which are classified as unmyelinated C fibers (red) and thinly myelinated Aδ fibers (orange); myelinated Aβ fibers (blue) transduce innocuous stimuli. The cell bodies of these peripheral sensory neurons are located in the dorsal root ganglia and trigeminal ganglia. Nociceptors innervate the epidermis and viscera, transducing noxious stimuli through the activation of several ion channels such as members of the TRP family. Finally, the information is transmitted to second order neurons located in the dorsal horn: C fibers and Aδ fibers mainly establish contact with neurons from layers I and II, while tactile Aβ fibers end mainly in layers III and V.
The Aδ-fibers are characterized by 2–6 μm diameters, are myelinated and present a relatively fast conduction speed between 5 and 30 m/s. The central branch of these fibers reaches the superficial laminae of the dorsal horn. These nociceptor fibers allow the conduction of cold, pressure and heat signals [5].
Remarkably, C-fibers are the ones to mainly establish nociceptive innervation. These are unmyelinated axons with a diameter of 0.4 to 1.2 μm and a conduction speed between 0.5 and 2 m/s. C-fibers innervate laminae I and II of the dorsal horn of the spinal cord and are activated in a polymodal fashion by mechanical, thermal (noxious cold or heat) and chemical stimuli [4, 5].
Diverse injurious stimuli detected by nociceptors are transformed into electrical activity, a process known as transduction where thermal, mechanical, or chemical signals are converted into ion flux through the activation of specialized ion channels [1]. In this way, noxious signals trigger fluctuations in the electrochemical gradient of nociceptors leading to membrane depolarization and subsequent action potentials, as a direct consequence of changes in the activation of ion channels expressed in the nociceptors [6]. Action potentials reach second-order neurons housed in the superficial laminae of the dorsal horn that project towards the spinothalamic and/or spinoparabrachial tracts contributing to the perception of noxious stimuli [2, 3, 7].
The study of the impact of sex steroids on the processing of noxious stimuli has been relevant to our understanding of the underlying differences between females and males. In the last decades, growing evidence related to sexual dimorphism in the perception of harmful signals and the development of pain conditions has been highlighted [8]. Several studies in humans using mechanical, chemical, electrical, or thermal stimuli have suggested that women have more sensitivity to some noxious stimuli than men [8, 9, 10, 11]. However, other contrasting reports have concluded that there is no difference in nociception between genders [12, 13]. Even though these differences are still not clear, the most reasonable explanation pinpoints sex steroids as the most relevant effectors. For example, some conditions such as headache, back and temporomandibular pain increase with pubertal development in girls, where the gonadal steroids produce crucial changes in women physiology [14]. Remarkably, differences in the perception of thermal stimuli between different sexes have also been reported, suggesting that women have greater sensitivity to heat than men [15]. In relation to this, an epidemiological study indicated that low testosterone concentrations in women are associated with an increase in the sensation of cold [16], clearly suggesting that steroids influence nociception.
2. Nociceptive TRP channels
The functional properties of nociceptors are critically regulated by the activity of ion channels expressed in their plasma membrane. Some of these channels belong to the Transient Receptor Potential (TRP) family that play vital roles as detectors of a wide range of biological signals. Some TRP channels are crucial receptors within nociceptors since they function as molecular machinery to transduce several noxious sensory stimuli [17].
In mammals, the TRP family consists of 27 members grouped into 6 subfamilies based on their sequence homology as follows: TRPC (canonical), TRPV (vanilloid), TRPM (melastatin), TRPP (polycystin), TRPML (mucolipin) and TRPA (ankyrin) subfamilies. A seventh group is the TRPN (no mechanoreceptor) subfamily which is not found in mammals, although it is found in the fruit fly, in C. elegans and zebrafish [18]. These receptors are nonselective cation channels that show an overall tetrameric structure, with each monomer being a six-pass transmembrane (S1-S6) protein with a hydrophilic re-entrant loop between the last two transmembrane segments that give rise to the ion conduction pore in the tetrameric arrangement, Figure 2. These channels have cytosolic N- and C- termini which are highly variable between the different members of the TRP subfamilies [19].
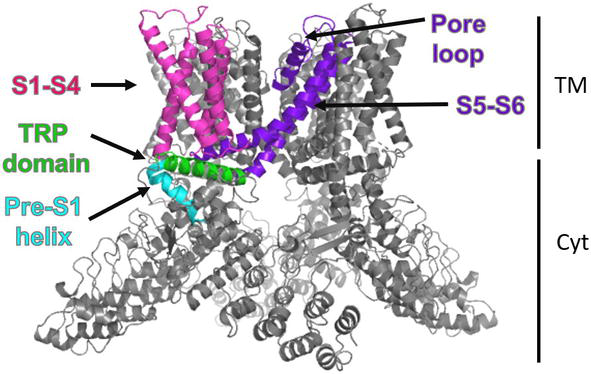
Overall 3D-structure of a TRP channel. Tetrameric assemble for TRP channels. The colored domains represent conserved domains in these channels: Transmembrane domain (S1-S6); the re-entrant loop forms the ion conduction pore; the pre-S1 helix joins the N-terminal to the S1; TRP box localized in the C-terminus. TM and Cyt: transmembrane and cytosolic regions. (TRPV1 3D- structure, PDB: 3J5P).
Some TRP channels are expressed in nociceptors where they display essential roles in the transduction of several noxious stimuli. Among these nociceptive TRP channels are TRPA1, TRPV1–4, TRPM2–3 and TRPM8 [17] which are activated by noxious thermal, chemical and mechanical signals and hence they are important pain-mediators, Table 1.
| Receptor | Temperature | Physical Stimulus | Natural Compounds | Endogenously-produced Compounds |
|---|---|---|---|---|
| TRPA1 | <17°C and noxious heat | Stretch | Allicin, Cinnamaldehyde, Allylisothiocyanate | Androstenedione Lysophosphatidic acid |
| TRPV1 | ≥42°C | Osmolality (hyper) Stretch | Capsaicin, Allicin, Resiniferatoxin, Tetrahydrocannabinol | N-arachidonoyl dopamine, Protons Lysophosphatidic acid Arachidonic acid and lipoxygenase-derived products, Anandamide |
| TRPV2 | ≥52°C | Osmolality (hypo), Stretch, Shear stress | Cannabidiol | _ |
| TRPV3 | 30–39°C | _ | Camphor, Gingerol, Zingerone, Carvacrol, Eugenol, Thymol | Farnesyl pyrophospahate |
| TRPV4 | 30–43°C | Osmolality (hypo), Shear stress | Bisandrographolide A | 5,6-epoxyeicosatrienoic acid |
| TRPM2 | >35°C | _ | _ | Adenosine diphosphate ribose, H2O2 |
| TRPM3 | ≥40°C | Osmolality (hypo) | — | Pregnenolone sulfate |
| TRPM8 | 10–28°C | Menthol, Eucalyptol | Artemin, Testosterone |
Nociceptive TRP channels and their main activators.
The TRPA1 channel, originally described as a cold sensor [20], is activated by several external compounds such as isothiocyanates contained in natural products such as wasabi, horseradish and mustard oil. Other natural TRPA1 activators are allicin and cinnamaldehyde, compounds found in garlic and cinnamon, respectively, and acrolein is also an exogenously-found compound that activates this ion channel [21, 22, 23, 24]. TRPA1’s activation by temperature in humans is still controversial, since some reports have described that TRPA1 is an insensitive cold-channel [23, 24], while several experiments have strengthened its role as a cold sensor [21, 25, 26, 27]. Furthermore, it has also been reported that TRPA1 channel has a redundant role as a noxious heat sensor in the mouse [28]. Indeed, this channel is also indirectly activated by inflammatory mediators like bradykinins and directly by lysophosphatidic acid (LPA, a compound released during injury tissue) and some reactive species produced as the 4-hydroxynonenal [24, 29, 30]. Thus, TRPA1 activation by these endogenous mediators produces inflammatory pain.
Among nociceptive TRP channels some members of the vanilloid receptors (i.e., TRPV1–4 channels) are also included. They are activated by warm (TRPV3-V4) to noxious heat (TRPV1-V2) and by several chemical compounds and osmotic changes.
The most studied channel of the vanilloid subfamily is the TRPV1 channel, originally described as the capsaicin receptor, since it is specifically activated by this pungent compound found in chili peppers [31]. This channel is also activated by compounds such as allicin, resiniferatoxin (a toxin found in the cactus-like plant Euphorbia resinifera) and by animal toxins such as the double-knot (a peptide toxin from the Earth Tiger tarantula), among others [31, 32, 33]. Additionally, several reports have demonstrated that TRPV1 activation is achieved by extracellular acid and intracellular basic pHs and heat (≥42°C) [31]. Furthermore, TRPV1 activation is produced by diverse compounds released during inflammation and tissue injury, for example, some arachidonic acid- and lipoxygenase-derived products, LPA and anandamide [34, 35, 36]. The high expression of the TRPV1 channel in small and medium-diameter neurons and its polymodal activation through several endogenous noxious stimuli, highlight it as a portal to pain; thus, the understanding of its regulation is a promising field to develop new strategies in the research field of pain and analgesia.
The vanilloid subfamily contains two less studied than TRPV1 members: TRPV2 and TRPV3 channels. The former is expressed in medium- and large DRG neurons, activated by temperatures with a threshold of 52°C [37] and by mechanical stretch and osmotic swelling [38]; exogenous compounds like some cannabinoids and endogenous lipids as lysophosphatidyl choline are also TRPV2 activators [39, 40]. Interestingly, the key role of TRPV2 as a noxious sensor is related to mechanical nociception in the somatosensory system, where it is required for the detection of noxious mechanical stimuli [41].
The TRPV3 channel is expressed in keratinocytes and co-expressed with TRPV1 channels in small-diameter sensory neurons. This channel is activated by temperatures between 30 and 39°C [42, 43] and camphor and cannabidiol are exogenous compounds that are also TRPV3 activators [44, 45]. Interestingly, it has been reported that polymorphisms found in the human TRPV2 and TRPV3 genes are associated to the susceptibility and severity to fibromyalgia, suggesting the role of these channels in chronic pain disorders [46].
The other vanilloid receptor playing a nociceptive role is the TRPV4 channel. This channel is activated by warm temperatures (~30–43°C) and hypotonicity [47, 48, 49]. Among the endogenous compounds that are TRPV4 activators are some derivates of arachidonic acid [50, 51]. Interestingly, this ion channel transduces osmotic stimuli inducing nociception and it is also an important mediator of the pruritogens effects of some compounds like serotonin [52, 53].
Finally, another TRP channels important to nociception are some members of the TRPM subfamily: TRPM2-M3 and TRPM8 channels. Although the TRPM2 channel is mainly expressed in immune cells, some expression has also been detected in neurons from DRG and TG [54, 55]. The TRPM2 channel is a warm thermo-sensor, since it is activated at 35°C [56]; other activators for this channel are adenosine 5′-diphosphoribose (ADPR) and H2O2[57, 58]. The activation of this channel through the generation of reactive species (ROS) implicates it as an important mediator of pain perception during stress oxidative conditions, that prevail in some conditions as fibromyalgia and neuropathy diabetic [59, 60].
The TRPM3 channel, initially identified in β-pancreatic cells [61] has an important role as a sensor of noxious heat, since it is activated by temperatures ≥40°C [62]. The activation of this channel can modulate glucose-induced insulin secretion and also nociception [61, 62]. Its role in the latter is due to high expression in small-diameter sensory neurons from DRG and TG [62] where it also serves as a sensor of noxious chemical stimuli to produce painful responses [62, 63]. Remarkably, TRPM3 together with TRPA1 and TRPV1 channels, form a redundant trio of sensors that mediates noxious heat transduction and provide protection against burn damage [28].
The group of nociceptive TRP channels also comprises the TRPM8 channel, a cold-transducer receptor (10–28°C) which is activated by natural compounds as menthol [64, 65]. This ion channel is predominantly expressed in a subpopulation of small-diameter sensory neurons lacking TRPV1 expression [64, 65]. TRPM8 overactivation is related to cold allodynia and hypersensitivity, pain caused by an innocuous cold stimulus and pain produce by enhanced sensitivity to cold stimulus, respectively [66, 67]. Similarly, to other TRP channels, some endogenous compounds such as artemin induces pain through TRPM8 activation [66]. Curiously, a steroid such as testosterone directly binds to this channel and induces its activation [68].
Clearly, the activation of these nociceptive channels is closely related to acute or chronic pain development and interestingly, some of these channels are targets for sex steroids actions, as we will describe here.
2.1 Estrogens and nociceptive TRP channels
Estrogens influence sexual differentiation in women and they have other essential functions in different biological processes as regulators of the cardiovascular system, metabolism, bone resorption and neuronal physiology [69]. These sex steroids comprise estrone (E1) and 17β estradiol (E2) and estriol (E3) [70]. Similarly, to other steroids, these estrogen’s actions are exerted by their interaction with specific nuclear receptors, the estrogen receptors alpha and beta (ERα and ERβ) that act as transcription factors to trigger gene regulation through a classical genomic pathway [71]. Furthermore, their actions also are through the activation of a membrane localized receptor, the G Protein-Coupled Estrogen Receptor 1 or GPER1 (previously named GPR30), establishing a non-classical pathway to regulate molecular targets [72, 73]. Several reports have suggested that estrogens influence some painful conditions like fibromyalgia, migraine, irritable bowel syndrome and temporomandibular disorder, all of which are conditions with major prevalence in women than in men [74, 75, 76, 77, 78].
Indeed, several experiments made in pain animal models have demonstrated the existence of sexual dimorphism in response to pain. For example, rats from a diabetic neuropathy model exhibited lower mechanical pain thresholds than males. This difference was abolished in gonadectomized animals, highlighting the sex-related differences in diabetic hypersensitivity [79].
These differences could be attributed to estrogen actions on different targets implicated in the transduction of noxious stimuli such as TRP channels, as will be described below.
A close relationship between estrogens and the TRPV1 channel has been demonstrated by several reports. Initially, it was stablished through behavioral assays that female rats exhibited increased capsaicin pain-related responses as compared to males. Furthermore, ovariectomized rats increased their pain threshold to capsaicin, which was reverted by replacement with E2 in these ovariectomized rats [80]. E2 effects on the TRPV1 nociception have also been evaluated during the rat estrous cycle, where the rats in the proestrus phase showed higher capsaicin-pain responses than the rats in estrus. This observation suggested that high E2 levels exhibited in the proestrus, positively regulate pain associated to TRPV1 activation [80]. These results have also been obtained evaluating the mechanical and thermal pain threshold during the mice estrous cycle, showing that mice in the proestrus displayed lower pain thresholds than the mice in estrus [81]. Remarkably, female TRPV1 knockout mice exhibited comparable mechanical and thermal pain thresholds during the proestrus and estrus [81], strongly suggesting that this channel has a pivotal role in the transduction of these noxious stimuli and that its activation is regulated by estrogen levels. A partial molecular mechanism through which estrogens regulate the capsaicin-evoked pain has been described. Initially, it was suggested that TRPV1 protein levels are downregulated in the DRG from the ERα and ERβ knockout mice-model [82], indicating that this ion channel is a molecular target for the effects of estrogen receptors, Figure 3. Furthermore, several reports have evidenced that TRPV1 mRNA levels are upregulated by E2. For example, primary cell cultures of rat TG or mice DRG neurons treated with E2 displayed increases in TRPV1 mRNA levels [81, 83]. This TRPV1 mRNA upregulation has also been observed in the hippocampus from ovariectomized rats treated with E2, heightening the allodynia displayed in a rat inflammatory temporomandibular model [84].
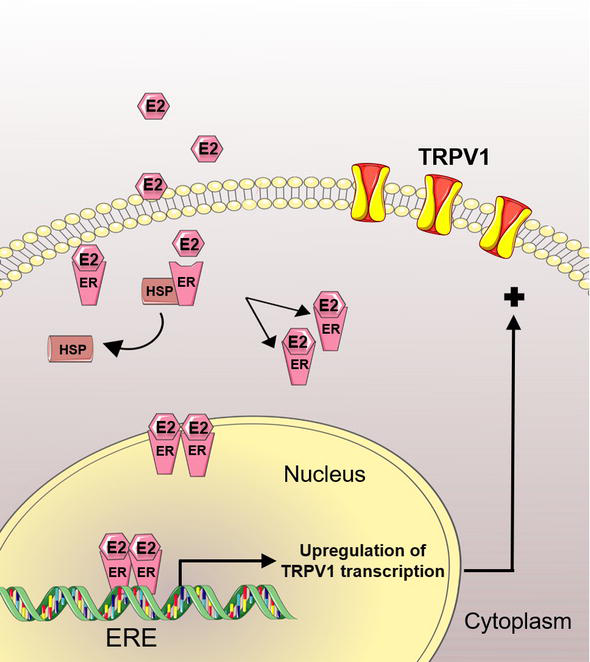
Schematic representation of the estrogen classical genomic effect in the regulation of TRPV1 gene expression. The inactive estrogen receptors (ER) localized in the cytoplasm form a protein complex with the heat shock protein (HSP). Internalized 17β estradiol (E2) interacts with the ER, activating and targeting them to the nucleus where they recognize the estrogen response element (ERE), a specific sequence located in the target genes (i.e., TRPV1 gene), resulting in positive regulation of TRPV1 expression.
The upregulation on TRPV1 gene expression has been also evidenced in synoviocytes, osteoclast precursors and endometrium [85, 86, 87]. Furthermore, experiments performed in cultures of human sensory neurons treated with E2 have shown increases in TRPV1 mRNA levels and similar results were obtained when the sensory neurons were incubated with a specific agonist of the ERβ [81, 88]. The E2-dependent effects in the aforementioned cell systems were observed after 24 h of treatment, suggesting that E2 achieves its actions through the classical genomic pathway. This possibility is strengthened by the identification of a putative estrogen response element (ERE) located in the promoter region of the mouse Trpv1 gene, implying that ERα could interact with this gene regulatory sequence to control TRPV1 gene transcription [89], Figure 3. These ERα bindings sites were also identified in the promoter sequences of the Trpv3–6 genes pointing out that estrogens are regulators of the transcription of these genes [89]. Additionally, it has been determined that different areas of the brains of mice in the proestrus (where the estrogens levels are high) display augmented mRNA levels for the Trpv2, v4 and v6 genes [89]. In contrast, this report also showed that TRPV1 and TRPV5 mRNA levels are downregulated in the brain from mice in the proestrus [89].
Additionally, negative E2 effects on TRPV1 activation have been evidenced in ovariectomized rats, which displayed reduced production of gonadal estrogens and augmented neuronal mitochondrial oxidative stress [90]. This was demonstrated using primary cultures of hippocampal and DRG neurons from these rats, determining that the current densities produced through TRPV1 activation are augmented. A similar effect was observed for the TRPA1 and TRPM2 currents [90]; consequently, overactivation of these channels caused rises in cytosolic Ca2+ and triggered apoptotic dead of these neurons. These effects were prevented in cultures of neurons derived from ovariectomized rats treated with E2 suggesting that this steroid could have neuroprotective actions, avoiding Ca2+ overload caused by the activation of some nociceptive TRP channels and preventing neuronal death [90].
Negative E2 actions on TRPV1 regulation have also been shown in primary cultures of rat DRG neurons treated with E2 or with an ERβ agonist, displaying decreases in capsaicin-evoked currents while TRPV1 protein levels remained unchanged [91]. This effect was lost using a non-permeable E2 (E2 conjugated to bovine serum albumin) and a slight increase in capsaicin-evoked currents was observed [91]. This result suggests that the role of E2 on TRPV1 activation requires steroid internalization in order to establish an interaction with ERβ and to negatively regulate this ion channel [91]. The discrete increase in TRPV1 activation using the non-permeable E2 is in agreement with a previous report which showed that E2 potentiated capsaicin-induced currents in rat DRG primary cultures [92]. Moreover, it was recently described that E2 potentiates TRPV1 activity in an estrogen receptor independent fashion, suggesting that TRPV1 could be an ionotropic receptor for E2 [93]. An interesting outcome from these experimental evidences is that the changes observed in TRPV1 expression are achieved using low E2 concentrations (10–100 nM) [81, 83, 88]; however, the effects on TRPV1 activation need supraphysiological concentrations of the steroid (1–100 μM) [93, 94]. This suggests that low E2 concentrations could modify TRPV1 expression through a classical genomic pathway, whereas a non-classical E2 effect on TRPV1 activation requires high E2 concentrations.
Non-classical E2 actions are produced faster than the genomic effects and are mediated by the interaction of this steroid with the GPER1 [72, 73]. The interaction of E2 with this receptor triggers different signaling pathways, such as those of protein kinase A (PKA) and C (PKC) [72, 95], which could produce TRPV1 phosphorylation. The phosphorylated state of the channel decreases its activation threshold to several stimuli, a process known as sensitization [96]. For instance, a phosphorylated TRPV1 channel can be activated at innocuous heat (i.e., 37°C) [96, 97]. TRPV1 phosphorylation also regulates the channel’s interaction with proteins like tubulin [98, 99], a protein association that is important to mediate mechanical gating of TRPV1 [100]. Interruption of the tubulin-TRPV1 complex through the TRPV1 phosphorylation on serine 800 (S800), decreases TRPV1 activation through osmotic stimuli [99, 100]. Interestingly, E2 through its association with GPER1, triggers the activation of PKCε signaling resulting in phosphorylation of TRPV1 in S800 and blocking the interaction of the channel with tubulin [99]; thus, E2 modifies microtubule-dependent TRPV1 mechanical pain sensitization.
The above data exemplify how TRPV1 is a target of classical and non-classical effects of estrogens, highlighting the estrogenic influence on nociception and pain mediated through the TRPV1 channel.
2.1.1 Estrogens upregulate TRPA1 and TRPV1 expression in endometriosis
Endometriosis is a debilitating chronic pelvic pain where the lining layer of the endometrium grows out of the uterus. This condition is highly estrogen-dependent and the lesion displays innervation of nociceptors co-expressing the TRPV1 and TRPA1 channels [101, 102]. Interestingly, mRNA levels for TRPA1 and TRPV1 are upregulated in the biopsies from women with endometriosis [88], thus it is feasible to consider that the overexpression of these channels could mediate the pain produced in this condition. The overexpression of these channels is likely to be through actions of estrogens, since they play a crucial role in the development of this anomalous pelvic condition. Particularly, it has been demonstrated that female rats treated with diethylstilbestrol, a non-selective estrogen receptor agonist, exhibited upregulation in the TRPV1 and TRPA1 mRNA levels [86] and this expression was found in endometrial cells. Moreover, cultured primary endometrial cells treated with this non-selective agonist also displayed increased TRPA1 and TRPV1 mRNA levels, whereas the treatment with E2 (which is a highly selective agonist of the ERα) only induced the upregulation of the TRPV1 mRNA levels [86]. The data suggests that these nociceptive TRP channels are regulated by the classical effects of estrogen receptors; additionally, these results showed the expression of these channels in non-neuronal cells from the rat endometria. TRPA1 and TRPV1 gene expression also has been demonstrated in human endometria from healthy women; furthermore, endometria samples from women suffering deep infiltrating endometriosis (one of the most severe forms of this condition) exhibit higher TRPV1 and TRPA1 expression, as compared to healthy women [103].
Additionally, it has been reported that hydroxylated estrogens (catechol estrogens) directly activate the TRPA1 channel [104]. This was evidenced using TRPA1-expressing HEK293 cells and whole-cell patch-clamp recordings that showed that 2-hydroxy-estrone (a catechol-estrogen) evoked currents through activation of TRPA1, similarly to another agonist for this channel, cinnamaldehyde. Thus, this hydroxylated estrogen acts as an endogenous agonist for this nociceptive channel [104]. Although, the physiological role of this activation has been demonstrated for insulin secretion in β-pancreatic cells, it would be interesting to explore if the excessive production of catechol-estrogens in endometriosis (as it has been previously reported [105]) affects the function of the TRPA1 channel expressed in endometrial cells and in the free nerve terminals surrounding this area. The expression of these channels in endometrial cells could represent a therapeutic alternative to relieve the main symptoms related to this painful condition; thus, it would be relevant to deepen the studies about the functional roles of TRP channels in these non-neuronal cells.
In addition to the above described evidence, the roles of TRPA1 and TRPV1 in endometriosis have been recently strengthened through the establishment of a mouse endometriosis model [106]. The endometriosis-like lesions induced in these animals displayed the presence of nerve fibers and inflammatory cells around the lesions. Remarkably, theses nerve fibers showed expression of TRPA1 and TRPV1 channels [106]. Furthermore, DRG neurons isolated from mice with induced endometriosis exhibited higher Ca2+ influx levels than neurons from sham animals, and this effect was through the activation of TRPA1 or TRPV1 channels [106]. The upregulation of these nociceptive channels could be directly associated with spontaneous abdominal pain observed in mice with endometriosis. Interestingly, the treatment of these mice with a synthetic androgen (danazol) or an aromatase inhibitor (letrozole), which are estrogen reducing agents, substantially relieved the spontaneously induced endometriosis pain [106]. These data reinforce the crucial role of estrogens in the development of endometriosis and highlight the importance of the TRPA1 and TRPV1 channels as molecular pain mediators in this debilitating form of abdominal pain.
2.2 Nociceptive TRP channels that are targeted by androgen actions
Androgens are steroids that influence the development of male characteristics and they are produced in the gonads (testicles and ovaries) and adrenal glands [107]. The main endogenously produced androgens are testosterone, androstenedione, dehydroepiandrosterone (DHEA) and dihydrotestosterone (DHT) [108]. Similarly, to other steroids, testosterone and DHT act as ligands for a specific protein, such as the androgen receptor (AR), while other androgens are weak ligands of the AR. This receptor regulates the expression of several target genes through a classical genomic pathway; furthermore, AR modifies cell physiology through non-classical and non-genomic effects [109].
Indeed, some androgens produce fast- non-genomic effects by interacting with other proteins like some TRP channels and modulating their function. For example, through whole-cell recording electrophysiological experiments, DHEA was shown to decrease capsaicin-dependent currents in dissociated DRG neurons [110]. DHEA effects seem to be direct and specific on TRPV1 channel activation, since with the use of a DHEA stereoisomer the inhibition of channel activation was no longer observed. Additionally, testosterone reduced the activation of TRPV1 to a lesser extent than DHEA [110]. This suggested the presence of a DHEA-binding site in the TRPV1 channel, which should be extracellularly localized, since the steroid was applied to the surface of the neurons during the electrophysiological recordings [110]. However, this remains undetermined and it is possible that DHEA has an antinociceptive role by inhibiting this channel.
The above data exemplified the rapid non-genomic effects of DHEA on TRPV1 activation. Additionally, testosterone seems to negatively regulate the expression of this channel, suggesting that TRPV1 is also a target for the classical actions of androgens. Supporting evidence for this was obtained in dissected TG from castrated male rats with orofacial myositis (induced by the Complete Freund’s adjuvant) [111]. The trigeminal TRPV1 mRNA and protein levels from this rat model were upregulated. Interestingly, TRPV1 expression is unaffected in the TG from castrated rats subjected to testosterone replacement [111], suggesting that testosterone prevents the increase in TRPV1 expression, although the molecular mechanism for this regulation is still unknown.
The TRPA1 channel is another molecular target for the non-classical effects of androgens. As a first approach, it was demonstrated that synthetic aromatase inhibitors evoked currents through TRPA1 channels in isolated DRG neurons; however, these compounds were unable to produce currents in DRG neurons from TRPA1 knockout mice, suggesting specific actions of the aromatase inhibitors on the activation of this channel [112]. Indeed, it was demonstrated that the aromatase inhibitors produced pain-like behavior in mice, a conduct not displayed in the TRPA1 knockout mice [112]. Since these compounds increase the levels of androstenedione in plasma, the effects of this androgen on TRPA1 activation have also been studied [113]. The results showed that androstenedione produces currents in mouse DRG neurons that are partially dependent on TRPA1 activation [113]. Furthermore, remanent androstenedione currents were abolished in the presence of the TRPV1 antagonist capsazepine, suggesting that this androgen could activate nociceptive TRPA1 and TRPV1 channels. Although, androstenedione is unable to cause acute pain-like behavior in mice, the animals exhibited mechanical allodynia 30 to 120 min after its intradermal injection into their paws, an effect that was abolished with a TRPA1 antagonist [113]. These data indicate that androstenedione could act as a pain sensitizing molecule leading to the activation of TRPA1.
Finally, the TRPM8 channel is also regulated by androgens. Interestingly, this channel is a target for the classical and nonclassical actions of testosterone. Initial evidence showed that TRPM8 is an androgen-responsive gene since testosterone treatment in some prostate cancer cell lines produced upregulation in the TRPM8 mRNA levels [114, 115]. TRPM8 expression also increases in the urogenital tract from orchiectomized rats supplemented with testosterone, suggesting that this steroid positively regulates TRPM8 gene expression [116]. Bioinformatic gene analysis showed that the TRPM8 gene contains putative sites for the androgen receptor interaction (androgen response elements, ARE) in its promoter sequence. Through chromatin-immunoprecipitation assays, it was confirmed that androgens receptors bind to these AREs located near the start transcription site of the TRPM8 gene [117]. These data indicated that androgens regulate TRPM8 gene expression in a classical genomic fashion.
The TRPM8 channel also acts as an ionotropic testosterone receptor due to direct interaction with this ion channel, resulting in its activation [68, 118]. Experiments have shown that the TRPM8 channel and the androgen receptor compete for testosterone and dihydrotestosterone because activation of TRPM8 by these androgens is lost when both receptors are co-expressed [68]. An extracellularly-located testosterone binding site in the TRPM8 channel has been suggested [68], and by using molecular docking simulation we have identified an additional intracellular testosterone pocket located between the firts intracellular loop (pre-S1) and the transmembrane segments 1 and 2 (S1-S2) of the TRPM8 protein [119]. This analysis has also shown that an H-bond is formed between testosterone and an arginine located in the TRP box, similar to the H-bond established between testosterone and an arginine in the aromatase enzyme [119]. The similarity in the testosterone pockets of the aromatase and the TRPM8 proteins could suggest that this is a functional binding site for TRPM8 through which activation by this steroid could be achieved in a non-classical and fast fashion. Interestingly, it has been demonstrated that topic testosterone application on humans produced cold- and pain-sensation; the women resulted more sensitized to the testosterone effects than men [68], suggesting a sexual dimorphism in testosterone-produced nociception through TRPM8 activation.
A third non-classical mechanism for TRPM8 regulation by testosterone has been demonstrated in DRG neurons where the androgen receptor and TRPM8 channels form a protein complex in the plasma membrane [120]. High testosterone concentrations (100 nM) inhibit TRPM8 channel activation and the androgen receptor is translocated to the nucleus [120], Figure 4. The evaluation of the TRPM8-mediated cold perception performed in castrated mice or rats revealed a decrease in their threshold for mild-cold perception; interestingly, testosterone supplementation in these animals inhibited cold perception (temperatures between 16 and 18°C) [120]. This result would suggest that testosterone confers endurance to TRPM8-mediated mild cold perception, which could explain the underlying molecular mechanism for the cold sensation experienced by women presenting lower levels of testosterone [16].
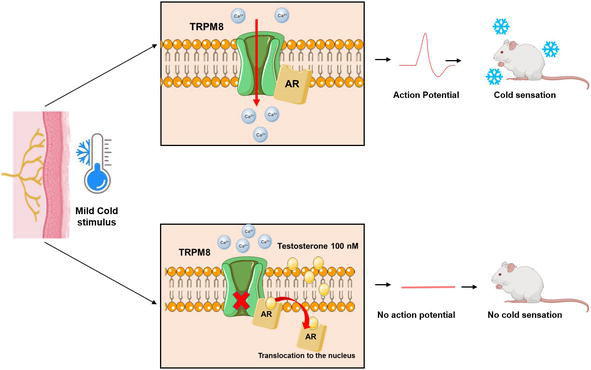
Testosterone regulates the TRPM8-mediated mild-cold sensitivity. The mild-cold stimulus is transduced through the TRPM8 channel located in the cell surface of DRG neurons where it forms a protein complex with the androgen receptor (AR) (upper panel). A high concentration of testosterone (100 nM) decreases activation of the TRPM8 channel and produces translocation of the AR to the nucleus. The physiological consequence of this is the loss of the mild cold sensation (lower panel).
2.3 Progesterone downregulates TRPV1 and TRPV4 channels expression
Progesterone is a steroid indispensable to the maintenance of pregnancy and it also plays important roles in the cardiovascular, immune, nervous systems and in the bones [121]. Progesterone is produced by the granulosa cells and the corpus luteum in the ovaries and, during pregnancy, the placenta is the major source of this steroid. Additionally, progesterone is produced in the nervous system, adrenal glands and testes [122]. The effects of progesterone are produced through nuclear receptors A and B (PRA/B), activating the classical genomic pathway to regulate gene expression. Furthermore, this steroid exerts a non-classical pathway through the activation of several membrane receptors (mPR) and other receptors as the progesterone receptor membrane components 1 and 2 (PGRMC1/2) and by the Sigma-1 Receptor (Sig-1R) [123, 124].
It has been demonstrated that progesterone regulates the expression of some TRP channels; for example, TRPV4 gene expression is negatively regulated by classical genomic progesterone effects [125]. Initial evidence for this was observed in epithelial and vascular cell lines and a recent report showed a correlation between TRPV4 expression and progesterone levels during the menstrual cycle, demonstrating that during the luteal phase when progesterone reaches its highest levels, TRPV4 expression is downregulated in human endometrial biopsies [126]. Furthermore, it has also been reported that TRPV4 expression is downregulated by a synthetic progestin (levonorgestrel) in a human Fallopian tubal epithelial cell line and mouse oviducts which could affect the transport of fertilized oocytes causing ectopic pregnancy [127].
Our group has determined that progesterone impacts on nociception by regulating the expression of TRPV1 channels [128]. Interestingly, this regulation is through a non-classical mechanism that requires an intermediate protein, the Sig-1R, which is a dynamic chaperone mainly localized in the endoplasmic reticulum and that can also be mobilized to the plasma membrane and nuclear envelope [129]. Progesterone decreases the levels of channels localized in the cell surface in HEK293 cells transfected with TRPV1 cDNA [128]. But how does progesterone produce this effect? Our results have shown that this is achieved by disrupting a protein complex formed between TRPV1 channels and the Sig-1R and by affecting the proper folding of TRPV1. Consequently, the channels are degraded by the proteasomal pathway and capsaicin-evoked currents are reduced [128]. The TRPV1 downregulation through progesterone actions was also observed in primary cultures of DRG neurons and we also demonstrated that during mice pregnancy when there are high progesterone levels, the threshold to capsaicin-dependent pain-like behaviors are augmented in comparison to non-pregnant mice, suggesting that progesterone confers protection to pain produced through activation of TRPV1 [128]. This work exemplified the antinociceptive actions that progesterone exerts through the decrease in the levels of TRPV1 channels localized in the plasma membrane of nociceptors and by the reduction of the transduction of harmful signals through this ion channel. Moreover, it has also been reported that progesterone disrupts the protein complex formed between Sig-1R and the TRPA1 and TRPM8 channels [130], suggesting that this steroid could impact the nociception through these nociceptive TRP channels.
3. Conclusion
Nociception is a neuronal process that protects against injurious stimuli that can be endogenously produced during inflammation or tissue injury processes. The transduction of these signals requires the activation of ion channels such as some members of the TRP family. In this chapter we have described how sex steroids influence nociception by regulating some of the TRP channels important for this neuronal process. Here we have described the data showing that estrogens and their nuclear receptors (ERα/β) positively regulate the expression of the TRPV1 and TRPA1 genes through a classical genomic pathway. The reports above discussed also show that progesterone and its nuclear receptors (PRA/B) negatively regulate TRPV4 gene expression.
We have also compiled and discussed the evidence about androstenedione’s and testosterone’s non-classical effects in the regulation of the activation of TRPA1 and TRPM8 ion channels directly. We have suggested a testosterone binding-pocket in the TRPM8 channel and we have also pinpointed an androstenedione binding-pocket in the C-terminal and pre-S1 regions of TRPV1 and TRPA1 channels [119].
These observations strengthen the notion that androgens act as direct molecular regulators of some nociceptive ion channels function, playing dual roles as nociceptive and antinociceptive molecules. For example, androstenedione has nociceptive actions on TRPA1 and TRPV1 activation and testosterone at low concentrations (picomolar range) can produce cold sensation through the TRPM8 channel, while high testosterone levels could improve resilience to cold sensation inhibiting the currents through TRPM8. Finally, the literature here reviewed shows that the nonclassical sex steroid effects are mediated by membrane receptors or even chaperones such as the Sig-1R, affecting signaling pathways and/or protein stability. This last finding exemplifies the antinociceptive effects of progesterone by which this hormone reduces the transduction of noxious stimuli mediated by TRPV1 channel activation. These data open a research field in which we will deepen our knowledge in the role of molecular sexual dimorphism and the interplay between nociceptive TRP channels and sex steroids.