
Role of transient receptor potential ankyrin 1 (TRPA1) on nociception caused by a murine model of breast carcinoma

By Amanda Spring de Almeida, Flávia Karine Rigo, Samira Dal-Toé De Prá, Alessandra Marcone Milioli, Gabriele Cheiran Pereira, Débora Denardin Lückemeyer, Caren Tatiane Antoniazzi, Sabrina Qader Kudsi, Daniel Mendes Pereira Ardisson Araújo, Sara Marchesan Oliveira, Juliano Ferreira, and Gabriela Trevisan
Excerpt from the article published in Pharmacological Research, Volume 152, February 2020, 104576, ISSN 1043-6618, DOI: https://doi.org/10.1016/j.phrs.2019.104576.
Editor’s Highlights
- Breast carcinoma affects women worldwide and is responsible for the onset of pain that can be persistent and intense, decreasing the patient’s quality of life.
- The blockade of TRPA1 by its antagonists shows the importance of new therapeutic strategies for the improvement of breast cancer pain.
- The activation of several ion channels, including acid-sensing ion channels (ASICs) and transient receptor potential vanilloid 1 (TRPV1), and TRP ankyrin 1 (TRPA1), have been described as involved in cancer pain induction.
- TRPA1 is involved in mechanical and cold hypersensitivity, and facial grimace criteria induced by tumor injection in mice.
Abstract
Breast carcinoma causes severe pain, which decreases the quality of life of patients. Current treatments produce adverse effects and have limited efficacy. Transient potential receptor ankyrin 1 (TRPA1) is related to the onset of cancer and neuropathic pain. The aim of this study was to evaluate the involvement of TRPA1 in a model of breast carcinoma. We injected 4T1 cells in the fourth caudal mammary fat pad of female BALB/c mice, and after 20 days we observed mechanical and cold allodynia and spontaneous nociception behavior (mouse grimace scale detection, MGS). TRPA1 selective antagonist (HC-030031 or A-967079) administration or intrathecal administration of TRPA1 antisense (AS) oligonucleotide was performed. The activity of NADPH oxidase, superoxide dismutase (SOD) and hydrogen peroxide (H2O2) levels were evaluated. The chemical hyperalgesia produced by a TRPA1 agonist (allyl isothiocyanate, AITC) was also detected. The administration of TRPA1 antagonists, TRPA1 AS, or antioxidant, transiently attenuated MGS, or mechanical and cold allodynia. Intraplantar injection of AITC also caused nociception. NADPH oxidase or SOD activity and H2O2 levels were increased in the sciatic nerve and hind paw skin samples. The 4T1 cells did not express TRPA1, and the use of HC-030031 or α-lipoic acid did not reduce the cytotoxic effect of a chemotherapeutic drug (paclitaxel). Thus, TRPA1 could be investigated as a target for breast carcinoma pain treatment.
Graphical abstract
1. Introduction
Despite the current advances in therapy and efforts to reduce incidence and mortality, breast carcinoma is still a significant public health concern worldwide [[1], [2], [3]]. Pain is one of the most feared and troublesome symptoms among breast carcinoma patients [4,5]. Although cancer pain can occur at any time during the disease, it frequently increases with cancer progression, and prevalence of pain ranges from 75 % to 90 % in patients with metastatic or advanced-stage cancer [4]. The search for more effective treatments for breast carcinoma, as well as for the treatment of pain due to the disease, emphasizes the importance of the morbidity of this pathology [6,7].
Despite the amount of information available on the treatment of cancer pain, a considerable proportion of cancer patients receive insufficient treatment [8]. Thus, unrelieved pain continues to be an essential concern [9]. At this time, opioids are the most effective analgesics available for the treatment of moderate to severe cancer pain. However, continuous opioid use can lead to several adverse effects, such as constipation and hyperalgesia [10,11]. Given that the analgesics used to treat cancer pain are ineffective and cause critical adverse effects, studies have been carried out to find innovative treatments [12].
The mechanisms associated with cancer pain are still uncertain, but the activation of several ion channels, including acid-sensing ion channels (ASICs) and transient receptor potential vanilloid 1 (TRPV1), and TRP ankyrin 1 (TRPA1), have been described as involved in cancer pain induction [13]. It has also been reported that activation or sensitization of nociceptors by protons or oxidant compounds leads to hypersensitivity in cancer pain [14,15]. The TRPA1 is a non-selective cation channelco-expressed with TRPV1 in peptidergic sensory fibers [16,17]. This receptor can be activated by natural irritants (including allyl isothiocyanate found in mustard oil) and oxidant compounds produced in the site of tissue injury (4-hydroxynonenal and hydrogen peroxide). Also, it can also be activated/sensitized by protons and inflammatory substances [18,19,70]. The role of TRPA1 has been investigated in neuropathic pain induced by several chemotherapeutic drugs, where it seems to be activated by oxidant compounds [20,21]. TRPA1 also appears to be essential to the maintenance of neuropathic pain induced by trauma or chemotherapeutic drugs [[20], [21], [22], [23], [24]]. Our research group recently showed that TRPA1 antagonism, or genetic deletion of the receptor, reduced the nociception observed in a model of metastatic melanoma in mice [13]. However, the involvement of TRPA1 in a model of breast carcinoma-induced nociception was not evaluated. It has also been reported that administration of 4T1 breast cancer cells to the mouse mammary gland leads to the development of bone metastasis, leading to mechanical and cold allodynia [25,26]. Thus, the current study was aimed at assessing the participation of the TRPA1 receptor in a model of breast carcinoma in mice.
…
3. Results
3.1. Antinociceptive effect of TRPA1 antagonists in a murine model of breast carcinoma
The intragastric administration of TRPA1 receptor antagonists, HC-030031 (300 mg kg−1) or A-967079 (100 mg kg−1), showed an anti-allodynic effect in a model of breast carcinoma in mice. HC-030031 and A-967079 compounds presented mechanical and cold anti-allodynic effects at 1 and 2 h after its administration (Fig. 1A–D). Moreover, treatment with TRPA1 receptor antagonists decreased MGS in this model of cancer pain at 1 h after its administration (Fig. 1E and F).
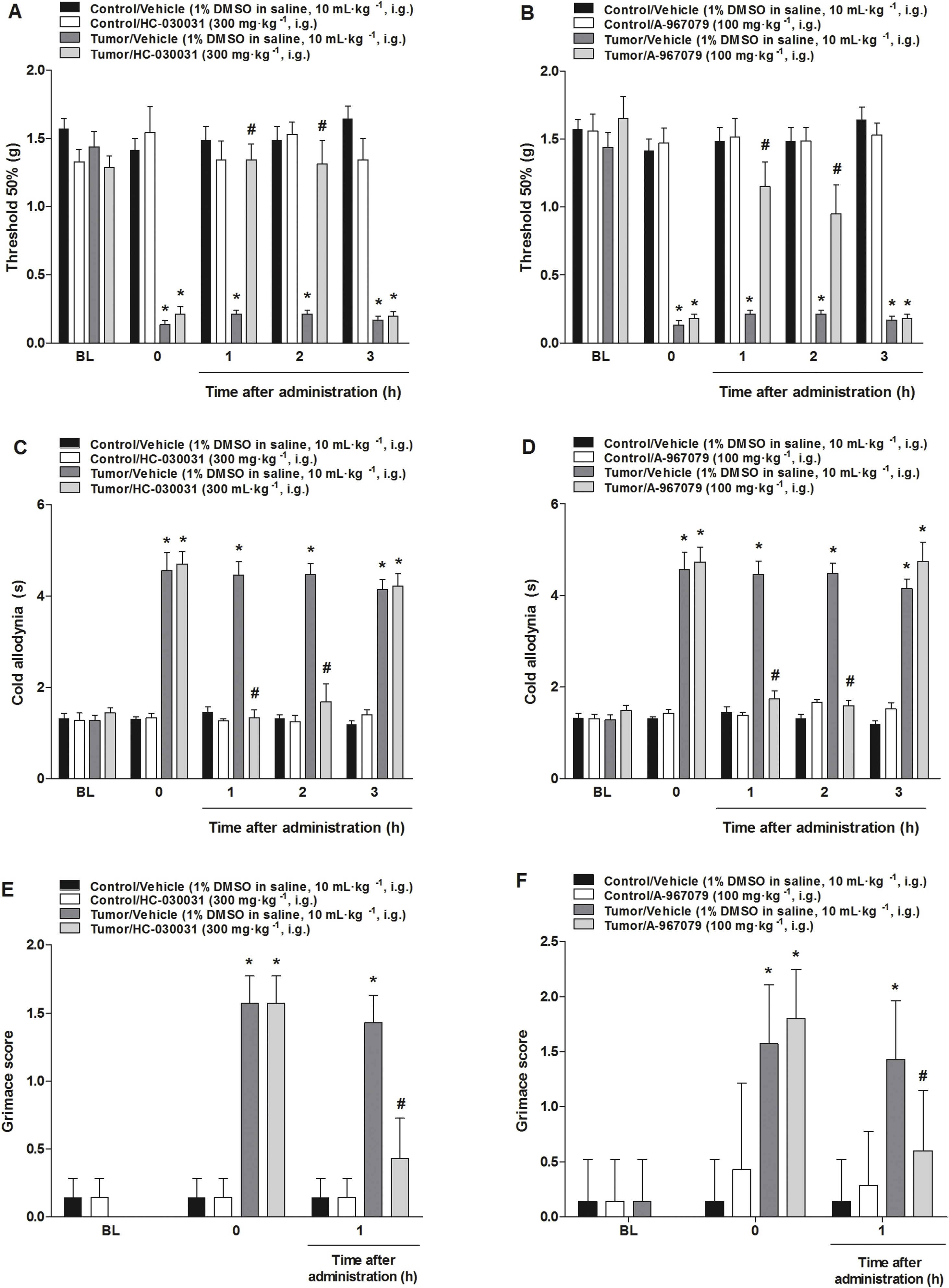
TRPA1 selective antagonist intragastric administration reduced mechanical and cold allodynia or diminished the mouse grimace scale (MGS) in a murine model of breast carcinoma.
(A, B) Mechanical and (C, D) cold allodynia, or (E, F) MGS alteration detected 20 days after tumor cells inoculation or vehicle (control group). Intragastric treatment with HC-030031 (300 mg kg−1) or A-967079 (100 mg kg−1) was evaluated on a time curve of 1, 2 and 3 h to the mechanical threshold and cold allodynia and during 1 h to MGS at day 20. Data are expressed as mean + S.E.M. (n = 7). *P < 0.05, when compared to the control group or baseline values and #P < 0.05 when compared to tumor/vehicle group (Two-way ANOVA followed by Bonferroni’s post hoc test).
3.2. Antinociceptive effect of a TRPA1 receptor antisense oligonucleotide injection in a murine model of breast carcinoma
TRPA1 AS oligonucleotide intrathecal administration induced an anti-allodynic effect for mechanical and cold allodynia, and also reduced MGS score (Fig. 2). TRPA1 MM oligonucleotide administration did not alter mechanical threshold, nociceptive response caused by acetone test, or MGS (Fig. 2).
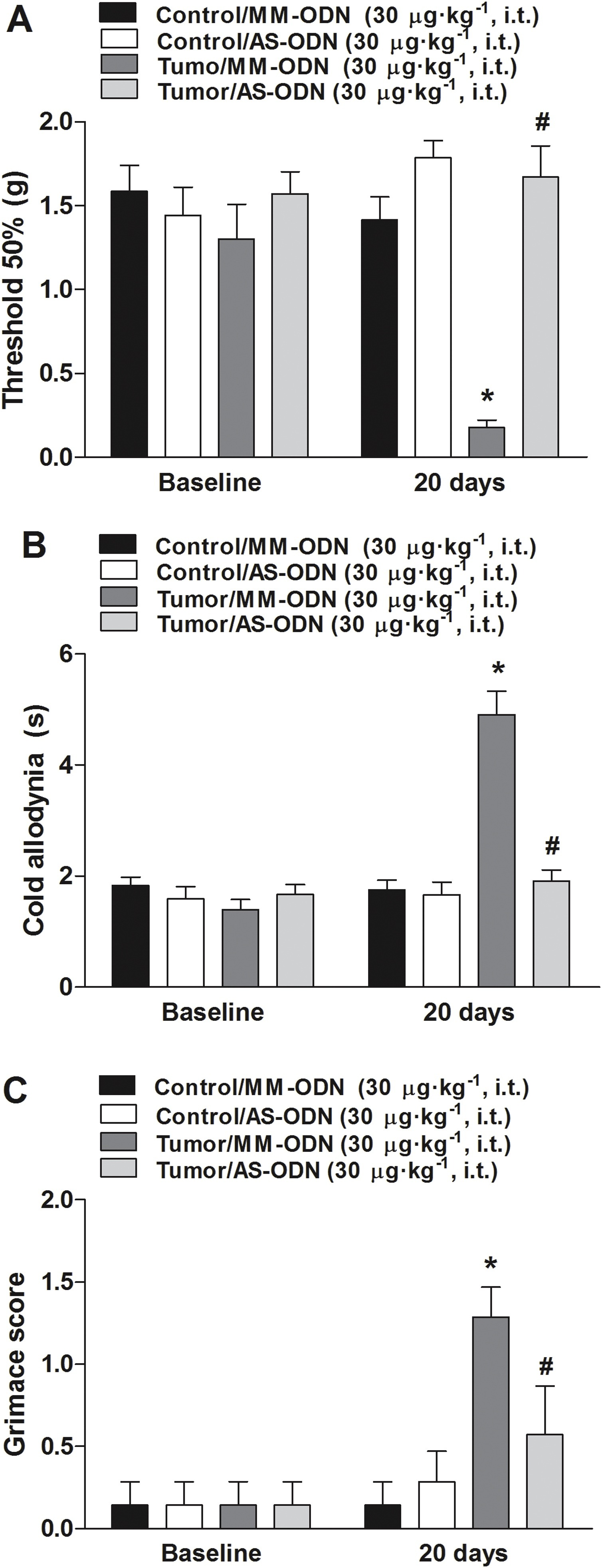
TRPA1 antisense (AS) intrathecal oligonucleotide administration reduced nociception observed in a murine model of breast carcinoma.
The AS antinociceptive effect was evaluated on (A) mechanical or (B) cold thermal stimulus and in (C) mouse grimace scales (MGS). The administration of mismatch (MM) oligonucleotide did not induce the antinociceptive effect. The nociceptive tests were performed 20 days after 4T1 cell inoculation. Baseline measures are graphically represented, and time 0 represents measures performed 20 days after tumor cell inoculation or PBS in mice. Data are expressed as mean + S.E.M. (n = 8). *P < 0.05, when compared to PBS+MM-treated group; #P < 0.05, when compared to tumor + MM-treated group (Two-way ANOVA followed by Bonferroni’s post hoc test).
3.3. The injection of 4T1 cells in the mammary gland in mice induced the production of TRPA1 agonists in sciatic nerve and paw skin
Tumor cell inoculation increased the H2O2 concentration (Fig. 3A and B), NADPH oxidase (Fig. 3D and E), and SOD activity (Fig. 3G and H) in both sciatic nerve and hind paw skin, and increased NADPH oxidase (Fig. 3F) when compared to the control group.
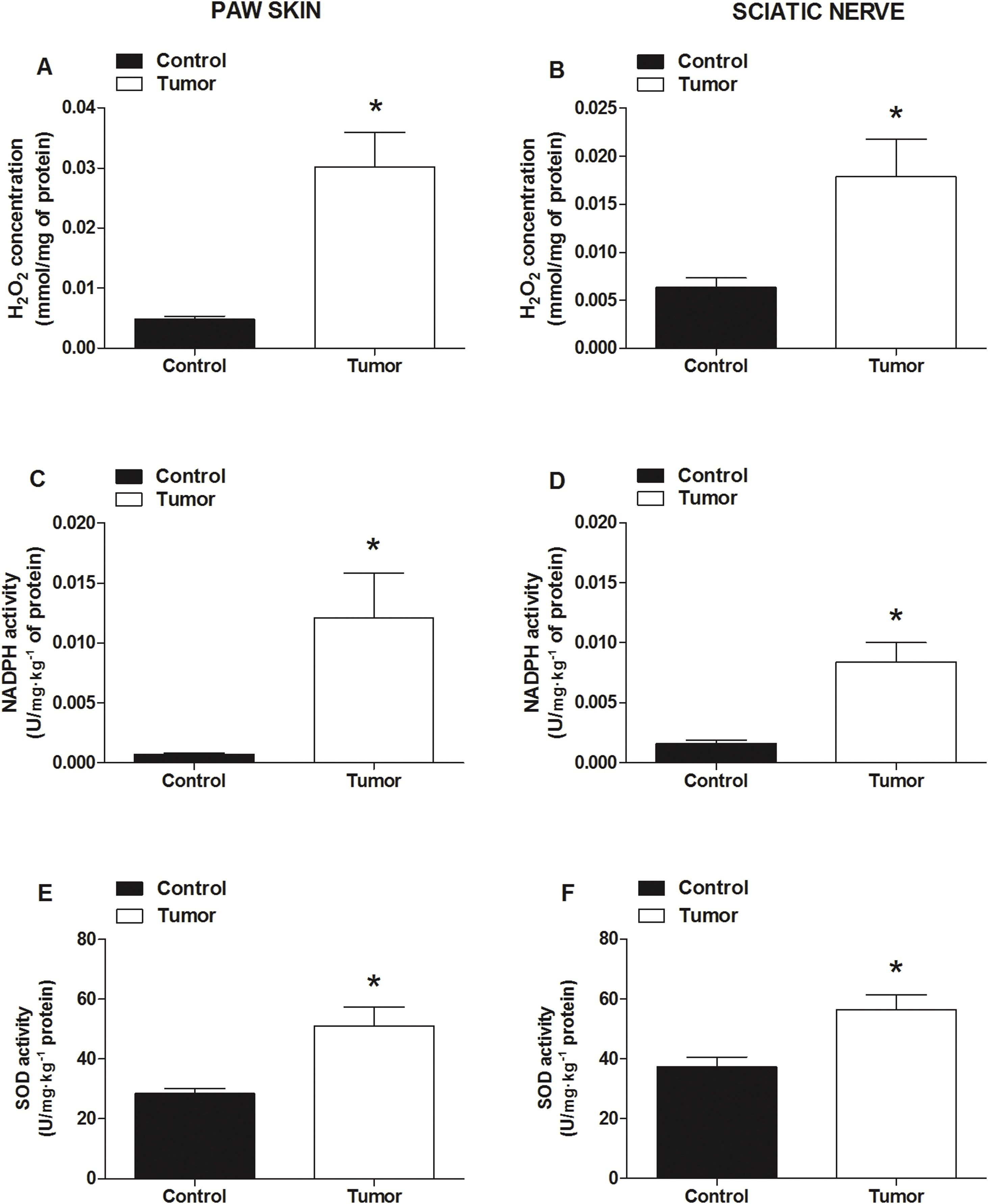
Administration of 4T1 tumor cells induced an increase in oxidative compound production and enzymatic activity in a murine model of breast carcinoma.
The oxidative status of cancer pain development was assessed through (A and B) H2O2 concentration, (C and D) NADPH oxidase and SOD (E and F) activity, in mouse sciatic nerve or skin paw tissues. The measurements were performed 20 days after 4T1 cell injection. Control group mice were injected with vehicle (PBS). Data are expressed as mean ± S.E.M. (n = 7). *P < 0.05, when compared to control (PBS) group (Student’s t-test).
3.4. The antioxidant α-lipoic acid caused an antinociceptive effect in a model of breast carcinoma in mice
The intragastric administration of an antioxidant, α-lipoic acid (100 mg kg−1), induced an anti-allodynic effect in a model of breast carcinoma in mice (Fig. 4A and B). α-lipoic acid presented a reduction in mechanical threshold 1 h after its administration, and this decrease was sustained for nociception time until 2 h after the antioxidant administration (Fig. 4A and B). Also, the antioxidant administration in animals that received only vehicle (PBS) paw injection did not alter mechanical threshold nor cold nociception measurements (Fig. 4). Moreover, treatment with α-lipoic acid decreased MGS in this model of cancer pain at 1 h after its administration (Fig. 4C).
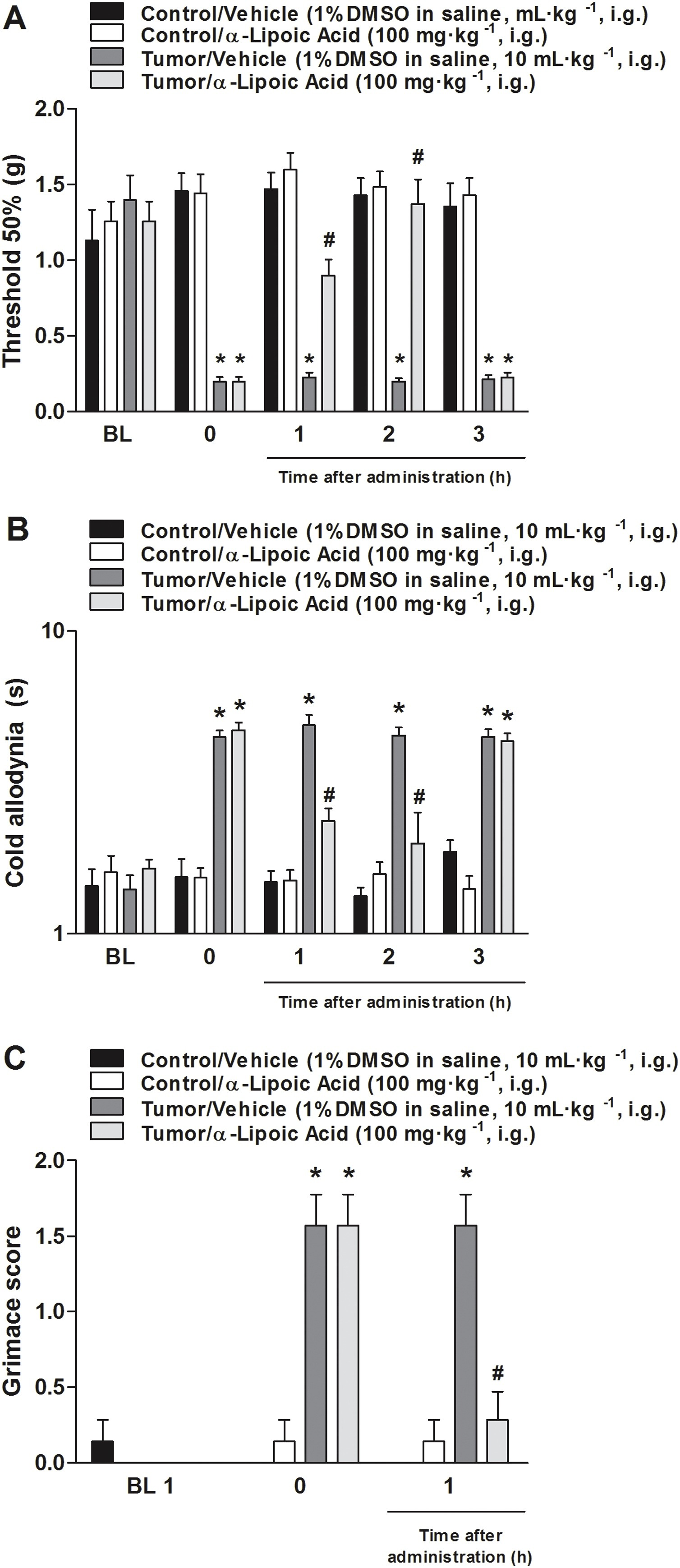
Antioxidant α-lipoic acid administration reduced mechanical and cold allodynia; this treatment also decreased the mouse grimace scale (MGS) in a murine model of breast carcinoma.
The α-lipoic acid (100 mg kg−1) effect was measured on (A) mechanical threshold, (B) cold allodynia, evaluated from 1 to 3 h after intragastric (i.g.) administration, and (C) for 1 h to MGS. The nociceptive tests were performed 20 days after 4T1 cell injection. Baseline measures are graphically represented, and time 0 represents measures performed 20 days after tumor cell inoculation or PBS injection in mice. Data are expressed as mean ± S.E.M. (n = 7). *P < 0.05, when compared to the vehicle+PBS-treated group; #P < 0.05, when compared to the tumor + PBS-treated group (Two-way ANOVA followed by Bonferroni’s post hoc test).
3.5. Intraplantar injection of HC-030031 or α-lipoic acid induced anti-allodynic effect and TRPA1 agonist (AITC) i.pl. injection caused chemical hyperalgesia in a murine model of breast carcinoma
The intraplantar injection of a TRPA1 antagonist or an antioxidant compound induced an antinociceptive effect in a model of breast carcinoma in mice (Fig. 5A and B). The antinociceptive effect of these compounds were local given that when HC-030031 or α-lipoic acid were injected in the contralateral paw, there was no antinociceptive activity in the measured hind paw (right side).
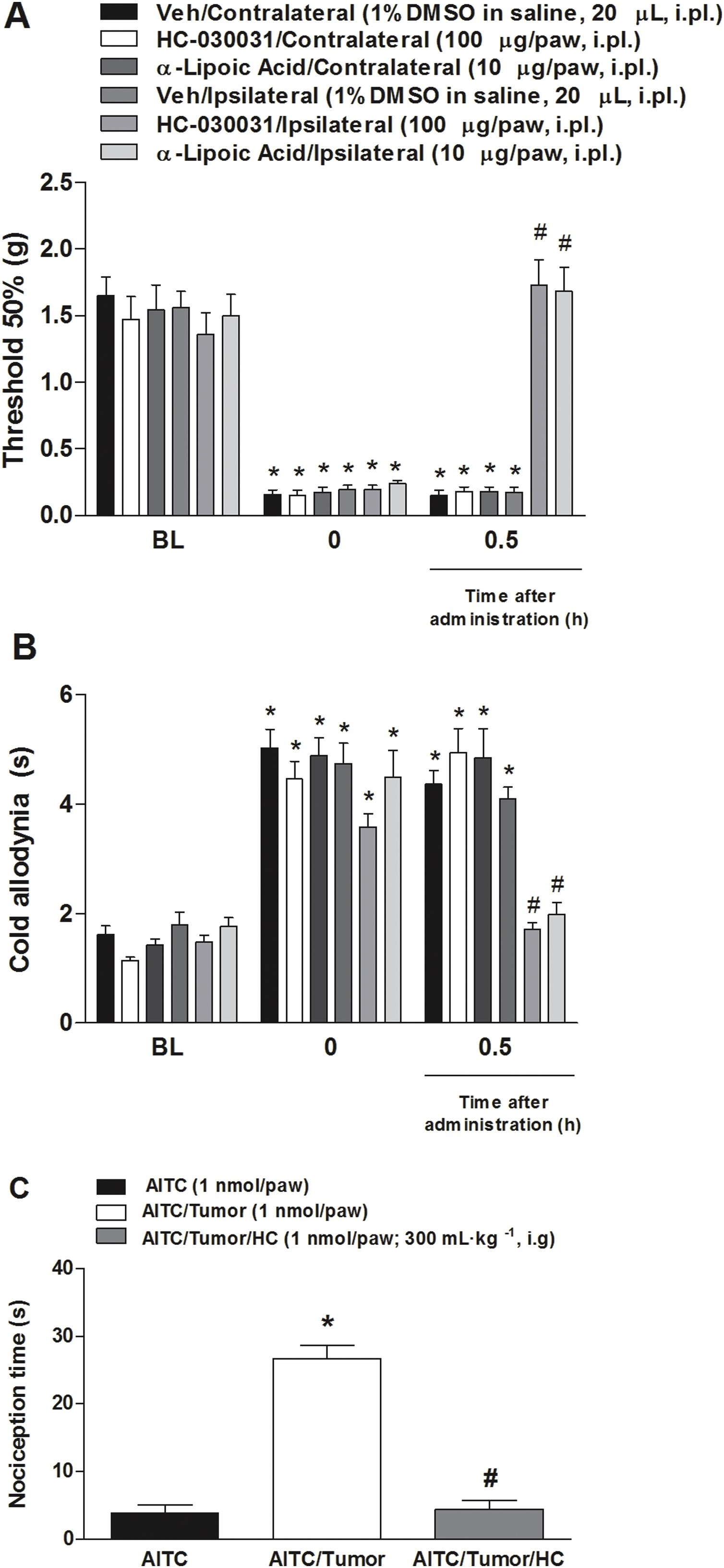
Local administration of α-lipoic acid and local administration of HC-030031 transiently reverse mechanical and cold hypersensitivity, and TRPA1 agonist administration (AITC) caused chemical hyperalgesia in a model of breast carcinoma in mice.
HC-030031 (100 μg/paw) or α-Lipoic acid (10 μg/paw) reduce the (A) mechanical and (B) cold allodynia induced by injection of 4T1 cells after 20 days in the paw ipsilateral (ipsi) to the injection. Values are mean + SEM of (n = 7), *P < 0.05. (C) The AITC (1 nmol) was administrated in the hind paw of mice and the nociception time was 5 min. Treatment with HC-030031 (300 mg kg−1, i.g.) was 1 h before AITC injection. Data are expressed as mean + S.E.M. (n = 7). *P < 0.001, when compared to AITC+tumor group; #P < 0.001, when compared to AITC + tumor+HC group (Two-way ANOVA followed by Bonferroni’s post hoc test in graph A and B; One-way ANOVA followed by Bonferroni’s post hoc test in graph C).
Moreover, intraplantar administration of the TRPA1 agonist (AITC, 1 nmol/paw) causes chemical hyperalgesia in a metastatic cancer pain model in mice (Fig. 5A). In mice treated 1 h before with the TRPA1 antagonist HC-030031 (300 mg kg−1), the spontaneous nociception behavior induced by chemical hyperalgesia was reduced (Fig. 5C).
3.6. HC-030031 and α-lipoic acid did not reduce the paclitaxel cytotoxic effect on 4T1 cells in vitro
The toxicity development of HC-030031 and α-lipoic acid was assessed in vitro using the 4T1 cell line through MTT assay, which revealed that HC-030031 and α-lipoic acid did not alter cell viability, regardless of the concentrations tested in comparison with the control group (Fig. 6A and B). On the contrary, 4T1 incubation with paclitaxel (PAC) showed a reduction in cell viability (30 and 100 μM), after 24 h of incubation when compared to control samples (Fig. 6C).
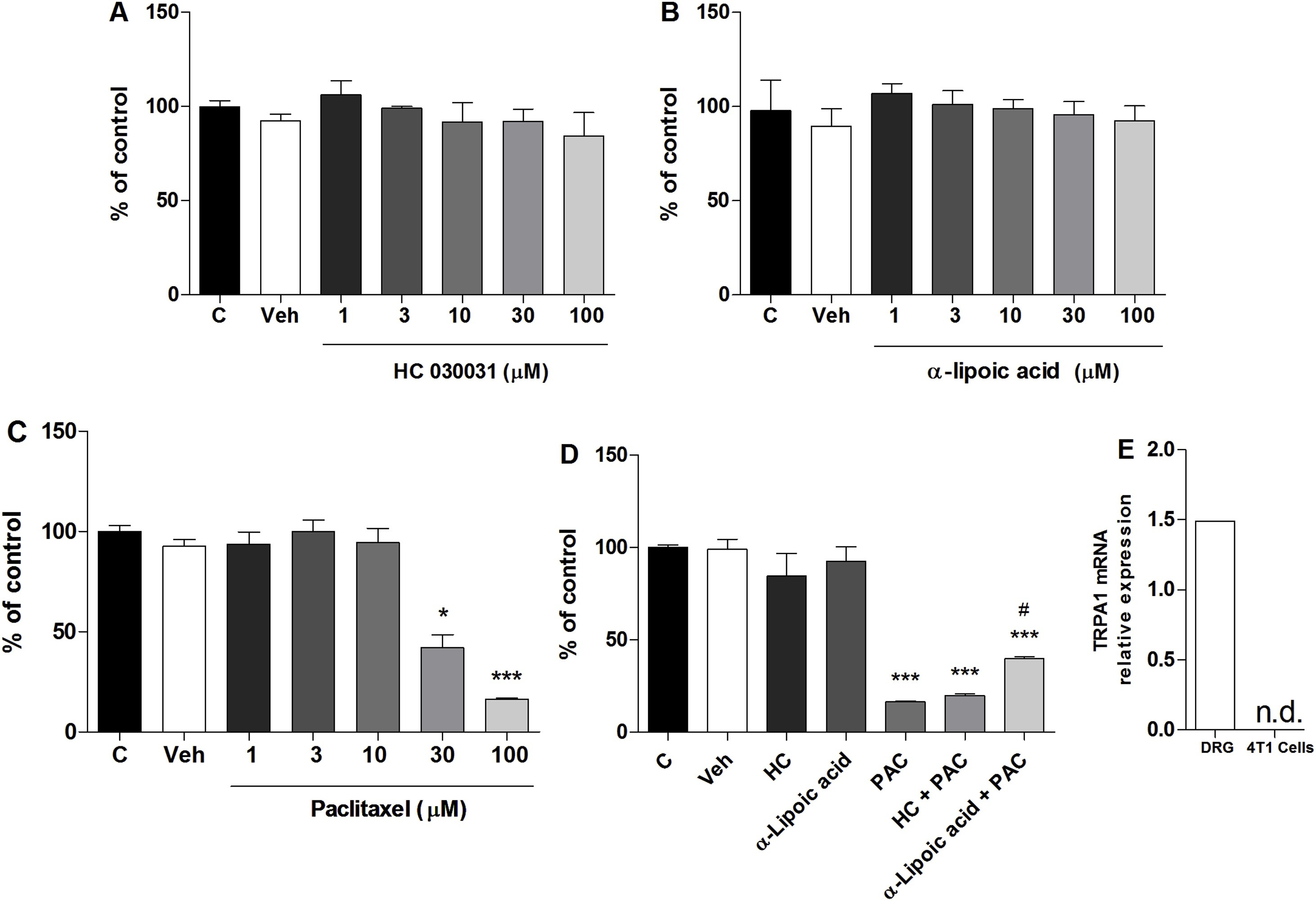
4T1 cell viability assessed in vitro by MTT assay and Reverse transcription PCR (RT-qPCR) for TRPA1 mRNA in DRG and 4T1 cells.
(A) HC-03003 (1–100 μM), (B) α-lipoic acid (1–100 μM), (C) paclitaxel (PAC, 1–100 μM). (D) HC-030031 (100 μM), α-lipoic acid (100 μM) associated with PAC (100 μM). (E) Control samples denote only DMEM culture medium. Control plus DMSO samples indicate DMEM culture medium plus DMSO 1 %. *P < 0.05, ***P < 0.001, when compared to control samples (One-way ANOVA followed by Bonferroni’s post hoc test). # P< 0.05 when compared PAC to α-lipoic acid + PAC (One-way ANOVA followed by Bonferroni’s post hoc test in graph A, B, C and D; Student’s t-test in graph E).
In the second set of experiments, the association of HC-030031 and α-lipoic acid with paclitaxel to cell viability was evaluated, to observe whether HC-030031 and α-lipoic acid interfered with the PAC chemotherapeutic effect. MTT assay showed that HC-030031 at the highest concentrations (100 μM), when associated with PAC highest concentration (100 μM; able to decrease cell viability, as described above) did not interfere with the chemotherapeutic action related to cell viability, but α-lipoic acid reduced PAC induced toxicity (Fig. 6D).
3.7. Reverse transcription PCR (RT-qPCR) and Sequence Read Archive (SRA) experiments showed no TRPA1 expression in the 4T1 cells
The 4T1 cells were examined for TRPA1 mRNA levels, to confirm the absence of TRPA1 receptors. There was no amplification for the 4T1 cell samples assessed by qRT-PCR, confirming the lack of TRPA1 expression in these cells, but we observed the TRPA1 mRNA presence in DRG samples (Fig. 6E). The qRT-PCR results were established through a second approach using the 4T1 cell RNA deep-sequencing in triplicate. We found no reads mapped against the trpa1 CDS for three independent transcriptome experiments. As an internal control, we used the gapdh gene, which presented more than 10,000 reads with 100 nt in size. A1 is not expressed in 4T1 cells grown in vitro.
4. Discussion
Breast carcinoma is a type of tumor that is highly capable of developing chronic pain [39]. Cancer pain is usually difficult to manage in the clinical setting, and several studies have been carried out to explore new mechanisms involved in this type of hypersensitivity, considering that unalleviated cancer pain expressively decreases the quality of life of patients [40]. Opioids are still the leading therapy used to reduce moderate to severe cancer pain [1,10]. Unfortunately, these medicines induce several adverse effects [41,42]. The TRPA1 channel has been reported to be involved in neuropathic pain induction and maintenance, especially neuropathic pain caused by chemotherapy, such as oxaliplatin, paclitaxel and bortezomib[20,21,23]. Consequently, if TRPA1 activation is also involved in cancer pain, treatment with TRPA1 antagonists could be used to manage cancer pain and chemotherapeutic-induced neuropathic pain, which are usually present in cancer patients [8,43]. In a recent study, we reported that TRPA1 is also involved in cancer pain in a model of metastatic melanoma in mice [13]. Our study was based on previous studies [25,26], which showed that, a few days after inoculation with 4T1 strain breast cancer cells in the mouse mammary gland, bone metastases develop. In addition, it has previously been shown [26] that the model of 4T1 cell inoculation in the mouse mammary gland, besides presenting bone metastases, causes the development of nociception observed as mechanical and cold allodynia. In this study we only measured the mechanical and cold allodynia in the right hind paw, but before when we characterize the model [26] we also detected the development of mechanical and cold allodynia in the left hind paw (data not show). In this model we observe that the onset of nociception occurs days after tumor cell inoculation, which is consistent with the onset of bone metastasis [25,26]. Metastatic cells travel to the bone, which is a favorable site for the development of metastases through the bloodstream, thereby inducing the production of inflammatory substances, which can activate receptors, thus leading to the onset of nociception. In addition, the tumor may compress or grow within a sensory nerve, leading to neuropathic pain [44,45]. It is important to note that the 4T1 cells used in the breast carcinoma model have already demonstrated high metastasis potential to bone and other tissues [25,26,46,47]. Thus, we could also relate the development of nociception with bone metastasis process caused by 4T1 cells inoculated in the mammary gland of mice.
We therefore explored the TRPA1 role in nociception observed in a murine model of breast carcinoma, using pharmacological and genetic tools. Our data showed that TRPA1 is involved in mechanical and cold hypersensitivity, and facial grimace criteria induced by tumor injection in mice. These findings were confirmed using TRPA1 antagonist (HC-030031 and A-967079) intragastric administration or by TRPA1 antisense oligonucleotide intrathecal injection. An increase in the production of H2O2 (a TRPA1 agonist), which could maintain TRPA1 activation/sensitization, was also found in this model. An antioxidant compound (α-lipoic acid) showed antinociceptive effect and reduced MGS in the current model, reinforcing the involvement of reactive substances in the maintenance of TRPA1 activation. Finally, all these findings indicate that TRPA1 participates in the development of breast cancer pain.
In the present study, we observed the presence of mechanical and cold allodynia 20 days after tumor cell injection, as well as an increase in MGS. These are interesting results, considering that spontaneous pain is often present in cancer patients [48], and spontaneous pain-related behaviors need to be better evaluated in preclinical pain studies [49]. The model used here could therefore be considered for the investigation of reflexive (using the von Frey test and acetone test) and non-reflexive (MGS) nociception measures in mice [26]. Moreover, we detected that i.g. administration of TRPA1 antagonists (HC-030031 and A-967079) reduced mechanical and cold hypersensitivity and reduced MGS in the mice. These antagonists were injected in commonly used doses to test the TRPA1 role in pain models in mice [20,21,32]. It has recently been described that TRPA1 plays an important role in the presence of pain in a metastatic melanoma model [13]. The TRPA1 participation in models of chemotherapeutic-induced neuropathic pain was also previously observed, which is a typical type of pain detected in patients with cancer [20,21,23,24]. Moreover, TRPA1 has been investigated to induce trigeminal and peripheral neuropathic pain after nerve injury [6,7,22,32], inflammatory pain[6,7,21], and migraine [50]. Thus, TRPA1 antagonists are described as potential analgesics to control chronic pain [50].
The results obtained using TRPA1 antagonists were confirmed using the TRPA1 antisense (AS) oligonucleotide intrathecal injection. TRPA1 AS injection abolished cancer pain-induced nociception and spontaneous pain-related behavior. These data are in line with those for the model of cancer pain caused by the intraplantar injection of melanoma cells [13]. Considering the data obtained by TRPA1 antagonist administration and TRPA1 antisense oligonucleotide injection, it seems that TRPA1 activation is relevant to nociception in this metastatic cancer pain model. The development of tolerance is an important adverse effect of opioid agonists. It would be interesting to evaluate this effect for TRPA1 antagonists in a model of cancer pain, given that opioids are the standard therapy for treating cancer pain. Previously, Wei et al. [69] showed that the tolerance induced by morphine administration was attenuated by a TRPA1 antagonist. Future studies should be conducted to determine whether repeated administration of TRPA1 antagonists develop tolerance, and a further study if TRPA1 blockage could hamper opioid-induce tolerance in a model of cancer pain.
TRPA1 is a known sensor for reactive compounds, such as acrolein [19], H2O2 [70] and 4‐hydroxy‐2‐nonenal (4-HNE) [18]. Thus, the TRPA1 channel could be activated or sensitized in this cancer pain model by an endogenous agonist, such as H2O2. It is well known that H2O2 is able to gate the TRPA1 channel [51], and its production has been associated with TRPA1 activation in neuropathic and inflammatory models of pain [22,32,52]. H2O2 levels were increased in mice 20 days after tumor inoculation, and NADPH oxidase and SOD activities were also augmented, a factor that could cause H2O2 increased production. It has been reported that reactive oxygen species could be involved in cancer pain development, but a potential receptor that could be gated by reactive compounds involved in cancer pain has not yet been reported [15]. It has been reported that administration of Walker 256 cells (rat mammary gland carcinoma cells) into the intratibial space in rats increases the levels of reactive oxygen species, such as H2O2, suggesting that ROS plays a role in cancer metastatic pain development [53]. The levels of 4-HNE have not been evaluated in a cancer pain model, but this mediator is relevant for the maintenance of TRPA1 activation in several models of pain [54]. Further supporting our findings, increasing levels of NADPH oxidase have already been shown in a murine breast tumor model [55]. Also, after the injection of B16-F10 cells into the hind paw [13], there was an increase in H2O2 production and NADPH oxidase activity in the paw skin.
The participation of reactive substances in this model was also detected using α-lipoic acid i.g. administration, which showed an antinociceptive effect and decreased spontaneous pain-like behavior in this metastatic cancer pain model. It was previously demonstrated that α-lipoic acid acute injection induced an antinociceptive effect in models of chemotherapeutic-induced neuropathic pain and neuropathic pain induced by nerve injury [21,22,32], revealing the role of reactive compounds in neuropathic pain. It has also been reported that treatment with HC-030031 and α-lipoic acid with local application decreased the nociception parameters in a persistent sensory neuropathy induced by a chemotherapy model in mice [21]. Here, we observed that local treatment with the TRPA1 antagonist and the antioxidant produces analgesia in a breast cancer pain model. The α-lipoic acid analgesic effect was previously described in patients, showing that this antioxidant has therapeutic capabilities for the treatment of pain-related disorders [56,57].
In a final set of experiments, we checked whether a TRPA1 antagonist or an antioxidant could reduce paclitaxel toxicity in 4T1 cells using the MTT assay. It was not tested before and is an interesting issue regarding paclitaxel is usually used to treat breast cancer [58,59]. It was found that HC-030031 did not alter paclitaxel chemotherapeutic effect, but α-lipoic acid altered its activity. Cell viability experiments testing drugs used in cancer treatment associated with other compounds have already been used in other studies [60,61]. These tests assist in research that seeks the possible synergism between substances already used and also to assess whether one substance interferes with the effect of another [60,61]. Some studies show the action of substances with analgesic effects without interfering in the chemotherapeutic action when using the cell viability test [[62], [63], [64]]. Duloxetine has been reported to improve painful chemotherapy-induced peripheral neuropathy in mice without interfering with the effect of oxaliplatin and paclitaxel [64]. In addition, another study demonstrated that the use of fingolimoid, used for the multiple sclerosis treatment, when used for the treatment of paclitaxel-induced peripheral neuropathy, has analgesic effect without altering the chemotherapeutic action in rats [62]. Still, it has been reported that cannabidiol also decreases the pain caused by paclitaxel in mice without modifying the action of this chemotherapeutic agent [63]. However, in our study, the association between α-lipoic acid and paclitaxel altered the chemotherapy action on the cell viability of breast cancer cells. This may be related to the fact that some antioxidant agents inhibit the formation of tumor cell free radicals and also inhibit the apoptosis of these cells, thus decreasing the sensitivity to chemotherapy [65]. However, it has been reported that the α-lipoic acid action in vivo, in rats, does not alter the action of paclitaxel [66]. Neither HC-030031 nor α-lipoic acid incubation alone altered cell viability. Moreover, 4T1 cells did not express the TRPA1 receptor, similar to that observed for B16-F10 cells [13].
Breast carcinoma affects women worldwide and is responsible for the onset of pain that can be persistent and intense, decreasing the patient’s quality of life. Currently, the best analgesic treatment is the use of opioids. However, these drugs cause numerous adverse effects [1,10,12,67]. Therefore, new strategies for the management of breast cancer pain should be sought out. We observed a high contribution of TRPA1 in the development of breast cancer pain, both in mechanical and cold allodynia and in spontaneous pain, which are the most problematic conditions of cancer pain [68]. The blockade of TRPA1 by its antagonists shows the importance of new therapeutic strategies for the improvement of breast cancer pain.