
Metabolomics and biochemical alterations caused by pleiotrophin in the 6-hydroxydopamine mouse model of Parkinson’s disease

By Esther Gramage, Jorge Sáiz, Rosalía Fernández-Calle, Yasmina B. Martín, María Uribarri, Marcel Ferrer-Alcón, Coral Barbas, and Gonzalo Herradón
Excerpt from the article published in Scientific Reports volume 12, Article number: 3577 (2022) DOI https://doi.org/10.1038/s41598-022-07419-6
Editor’s Highlights
- Parkinson’s disease (PD) is a neurodegenerative disorder that is characterized by the progressive loss of dopaminergic neurons in the substantia nigra (SN) and dopaminergic terminals of the striatum.
- Pleiotrophin (PTN) is one neurotrophic factor recently shown to be an important regulator of neuroinflammation in different pathological contexts.
- Pleiotrophin is a survival factor for dopaminergic neurons and promotes tyrosine hydroxylase (TH) expression.
- Striatal gliosis is a hallmark of PD and, in general, of the neuroimmune response triggered by neurotoxins such as 6-OHDA.
- The protective effects of PTN on dopaminergic neurons are paralleled by a regulation of potential glial responses after the administration of a low dose of 6-OHDA.
- Entire groups of phospholipids tend to vary together is very significant because α-Synuclein is a lipid-binding protein that interacts with phospholipids and fatty acids.
- Phospholipids are associated to neurodegeneration and data presented here support the increasingly recognized “lipid cascade” in PD.
Abstract
Pleiotrophin (PTN) is a cytokine involved in nerve tissue repair processes, neuroinflammation and neuronal survival. PTN expression levels are upregulated in the nigrostriatal pathway of Parkinson’s Disease (PD) patients. We aimed to characterize the dopaminergic injury and glial responses in the nigrostriatal pathway of mice with transgenic Ptn overexpression in the brain (Ptn-Tg) after intrastriatal injection of the catecholaminergic toxic 6-hydroxydopamine (6-OHDA) at a low dose (5 µg). Ten days after surgery, the injection of 6-OHDA induced a significant decrease of the number of tyrosine hydroxylase (TH)-positive neurons in the substantia nigra and of the striatal TH contents in Wild type (Wt) mice. In contrast, these effects of 6-OHDA were absent in Ptn-Tg mice. When the striatal Iba1 and GFAP immunoreactivity was studied, no statistical differences were found between vehicle-injected Wt and Ptn-Tg mice. Furthermore, 6-OHDA did not cause robust glial responses neither on Wt or Ptn-Tg mice 10 days after injections. In metabolomics studies, we detected interesting metabolites that significantly discriminate the more injured 6-OHDA-injected Wt striatum and the more protected 6-OHDA-injected Ptn-Tg striatum. Particularly, we detected groups of metabolites, mostly corresponding to phospholipids, whose trends were opposite in both groups. In summary, the data confirm lower 6-OHDA-induced decreases of TH contents in the nigrostriatal pathway of Ptn-Tg mice, suggesting a neuroprotective effect of brain PTN overexpression in this mouse model of PD. New lipid-related PD drug candidates emerge from this study and the data presented here support the increasingly recognized “lipid cascade” in PD.
Introduction
Parkinson’s disease (PD) is a neurodegenerative disorder that is characterized by the progressive loss of dopaminergic neurons in the substantia nigra (SN) and dopaminergic terminals of the striatum. The underlying cause of this neuronal loss is not completely understood. Neuroinflammation, characterized by excessive microgliosis and astrogliosis, has been implicated in the pathophysiology of PD, being critical to PD progression1,2. Thus, identification of the molecular mechanisms involved in dopaminergic cell survival and modulation of the neuroimmune response represents an opportunity to discover novel therapeutic targets and/or biomarkers in PD. Pleiotrophin (PTN) is one neurotrophic factor recently shown to be an important regulator of neuroinflammation in different pathological contexts3.
Pleiotrophin is a survival factor for dopaminergic neurons and promotes tyrosine hydroxylase (TH) expression4. PTN is also a survival factor for the catecholaminergic PC12 cell line5. Genetic inactivation of Ptn exacerbates amphetamine-induced dopaminergic injury in the nigrostriatal pathway6, including the unexpected loss of 20% of dopaminergic neurons in the SN that is not observed in amphetamine-treated Wild type (Wt) mice. Interestingly, amphetamine- and cocaine-induced alterations in the striatal phosphoproteome of Ptn−/−mice were similar to those found in PD7,8,9, suggesting that PTN may protect against neuronal injury in different brain disorders. PTN is one of the factors upregulated after treatment with levodopa in the denervated striatum of parkinsonian rats10,11, suggesting a role for this cytokine in the neurotrophic mechanisms and plasticity triggered by treatment with levodopa. Interestingly, PTN is upregulated in the dorsolateral striatum in the 6-hydroxydopamine (6-OHDA) rat model of PD, suggesting neurotrophic actions of this cytokine in this model12. Supporting this, PTN is involved in the cAMP-dependent enhancement of the differentiation of dopaminergic neurons in cultures13, prevents amphetamine- and cocaine-induced toxicity in PC12 cells5,14 and exerts trophic effects on donor cells after neural transplantation in vivo to achieve functional recovery of nigrostriatal pathways in animal models of PD15. Furthermore, striatal and/or nigral PTN over-expression provides neuroprotection against the dopaminergic toxic 6-OHDA in rats16,17.
The proven neuroprotective effects of PTN in different models are interesting because Ptnexpression is upregulated in the brain in different pathologies such as PD, Alzheimer’s disease (AD), ischemia, and after administration of different drugs of abuse including amphetamine and alcohol3. In PD patients, PTN is upregulated in the degenerating substantia nigra, apparently in dopaminergic neurons with downregulated expression of TH18. Recently, loss of pericytes in PD, AD, amyotrophic lateral sclerosis (ALS), human immunodeficiency virus (HIV)-associated neurocognitive disorder and Huntington’s disease has been linked to PTN depletion causing neuronal loss19. These different disorders are characterized by overt neuroinflammation and PTN has been recently found to modulate the neuroimmune response in different pathological contexts. For instance, transgenic Ptn overexpression in the brain potentiates striatal astrocytosis induced by acute administrations of amphetamine20. Ptnoverexpression also potentiates microglial activation and increases of pro-inflammatory cytokines in the brain induced by an acute administration of lipopolysaccharide (LPS)21.
The evidence summarized here suggests that PTN potentiates the acute neuroimmune response induced by different stimuli, including microglial and astrocytic responses, which is necessary and critical for host defence22. However, persistent and/or over-activation of microglia is deleterious. Thus, one aim of the present work was to shed some light on the possible role of PTN in the subchronic nigrostriatal neuroinflammation associated with the 6-OHDA mouse model of PD. In addition, understanding altered metabolic pathways and metabolites involved in the development and progression of disease provides a better knowledge of the underlying related biological alterations. This is important for diseases like PD, for which the biomolecular causes are still unclear and, of particular interest in early stages of the disease when neuroprotective strategies are expected to be more successful. In these regards, metabolomics has become an important tool able to provide useful insights into unknown biochemical mechanisms and possible biomarkers for various disorders. For that reason, we aimed to confirm if Ptn overexpression in the mouse brain prevents dopaminergic injury in an 6-OHDA model with a partial lesion and how PTN modifies biochemical cascades, using an untargeted metabolomic approach.
…
Results
Absence of 6-OHDA-induced loss of TH contents in the nigrostriatal pathway of Ptn-Tg mice
Before sacrifice, animals were tested for alterations in motor functions by means of rotarod and cylinder tests. We did not observe any changes in motor functions caused by the treatment or the genotype (Supplementary Fig. 1). To test the possibility that Ptn-Tg mice may be protected against 6-OHDA-induced neurotoxicity in the SN, we assessed the number of TH+ cells in this area of vehicle- and 6-OHDA-treated Ptn-Tg and Wt mice. ANOVA revealed a significant effect of the treatment (F(1,14) = 12.17; P = 0.0036). Post-hoc comparisons revealed that vehicle-treated Ptn-Tg mice did not show significant differences in the number of TH+ cells compared to Wt mice (Fig. 1, P = 0.25). We found a significant decrease in the number of TH+ cells in the substantia nigra of Wt mice treated with 6-OHDA compared to vehicle-treated mice (Fig. 1, P = 0.0106). In contrast, we found that 6-OHDA did not cause a significant change in the number of TH+ cells of Ptn-Tg mice compared with vehicle-treated animals (Fig. 1, P = 0.64). The data suggest that the severity of TH+ cell loss in the SN induced by 6-OHDA is limited by PTN.
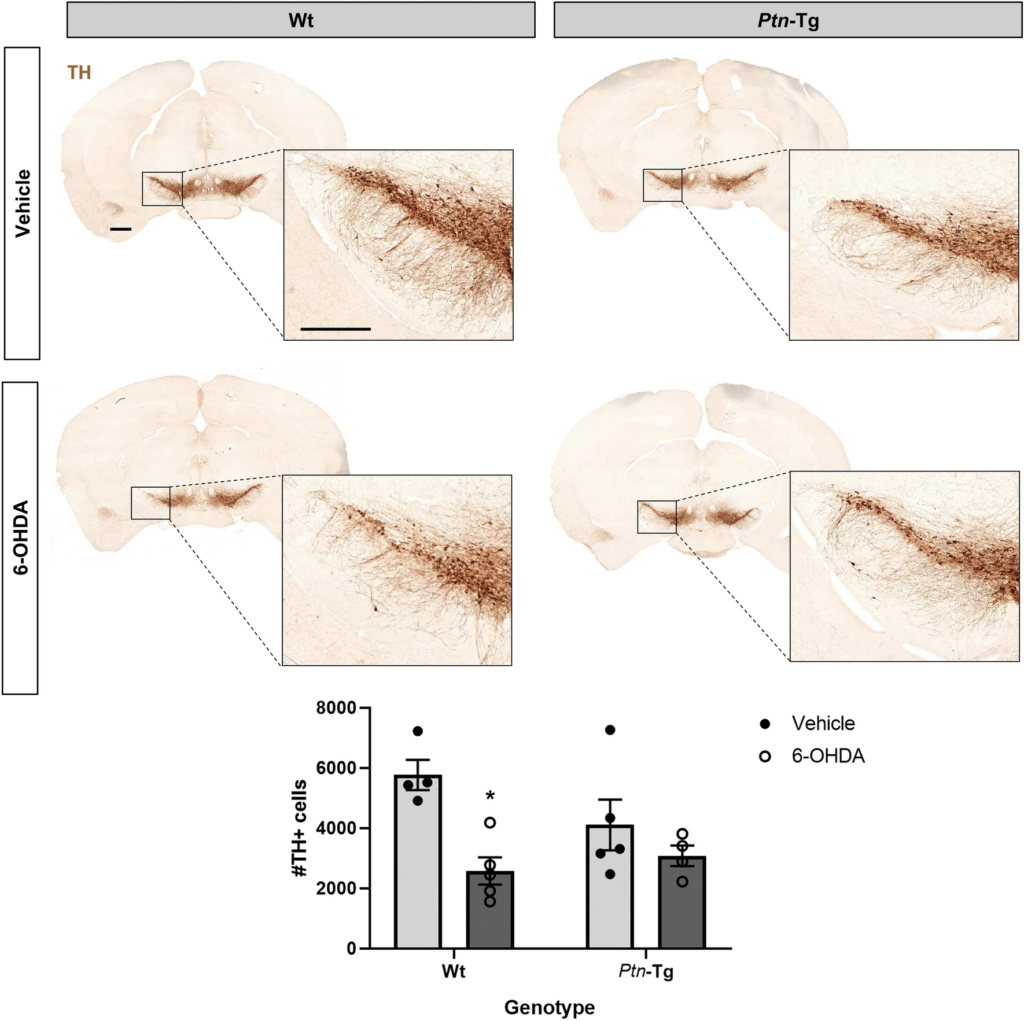
6-OHDA induces loss of TH+ neurons in the substantia nigra of Wt mice. Photomicrographs of substantia nigra of TH-immunostained sections from mice sacrified 10 days after vehicle or 6-OHDA striatal injection. Graph shows the number of TH+ cells in the substantia nigra pars compacta counted by stereology. Data are represented as mean ± S.E.M. *P < 0.05 vs. Wt-Vehicle. Scale bar = 500 μm (magnification image
We also analyzed in immunohistochemistry studies TH expression in the striatum of Ptn-Tg and Wt mice treated with 6-OHDA or vehicle (control). ANOVA revealed a significant effect of the genotype (F(1,15) = 32.37; P < 0.0001), a significant effect of the treatment (F(1,15) = 27.15; P < 0.0001) and a significant interaction genotype x treatment (F(1,15) = 24.65; P = 0.0002). Post-hoc analysis did not detect significant differences in the levels of TH in the striatum of vehicle-treated Ptn-Tg and Wt mice (Fig. 2, P = 0.99), suggesting that overexpression of PTN is not a key factor for TH expression in the mouse striatum in normal condition. We found that 6-OHDA caused a significant depletion of TH contents in the striatum of Wt mice compared with vehicle-treated Wt mice (Fig. 2, P < 0.0001). Interestingly, we did not observe a decrease of TH levels in the striatum of Ptn-Tg mice treated with 6-OHDA (Fig. 2, P = 0.96). The data confirm that 6-OHDA produces degeneration of dopaminergic terminals in the striatum of Wt mice, effect that was absent in Ptn-Tg mice. Overall, the data suggest that PTN modulates the severity of 6-OHDA-induced neurotoxicity in the mouse nigrostriatal pathway.
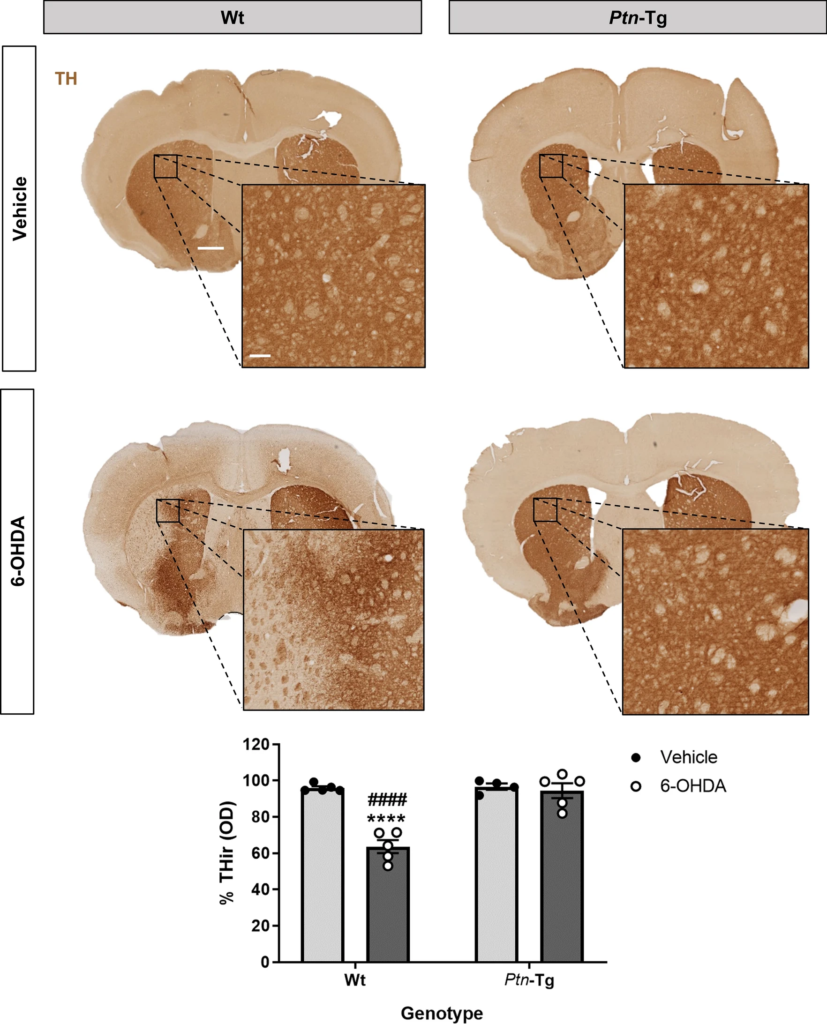
6-OHDA-induced loss of TH expression in striatum is prevented in Ptn-Tg mice. Photomicrographs illustrate that PTN overexpression prevents 6-OHDA-induced dopaminergic damage. TH-immunostained striatal sections of mice, 10 days after vehicle or 6-OHDA striatal injection. Graph shows the proportional stained area of TH-ir (TH-immunoreactivity) in the striatum. Data are represented as mean ± S.E.M. OD: optical density. ****P < 0.0001 vs. Wt-Vehicle. ####P < 0.0001 vs. Ptn-Tg-6-OHDA. Scale bar = 500 μm (magnification image scale bar = 100 μm).
6-OHDA-induced glial responses in Wt and Ptn-Tg mice
Taking together, the data presented here support a role of PTN against 6-OHDA-induced neurotoxicity in nigrostriatal dopaminergic circuits. Since PTN is a novel modulator of glial activation and neuroinflammation in different contexts3,21,36,37, we aimed to study the modulatory role of PTN overexpression on 6-OHDA-induced striatal glial responses. We tested the astrocytic and microglial response in striatal sections of vehicle- and 6-OHDA-treated Wt and Ptn-Tg mice. The ANOVA of the immunohistochemistry studies of the microglial marker Iba1 did not show treatment-related differences but revealed that the number of Iba1+ cells and marked area tended to be different between genotypes (Iba1+ cells, F (1,18) = 3.866, P = 0.0649; marked area, F (1,15) = 3.971, P = 0.0648) (Fig. 3a), whereas significant differences between genotypes were observed in the case of Iba1 cell size (F (1,18) = 5.577, P = 0.0297). The data reflects higher values of these three parameters in Wt mice; however, post-hoc analysis of Iba1 cell size did not reveal significant differences between individual groups. The ANOVA of the data from the immunohistochemistry studies for GFAP, an astrocyte-specific intermediate filament protein38, did not reveal treatment-related differences nor differences between genotypes (GFAP+ cells, F (1,14) = 2.519, P = 0.1348; marked area, F (1,13) = 3.286, P = 0.0930; cell size, F (1,14) = 2.414, P = 0.1426) (Fig. 3b).
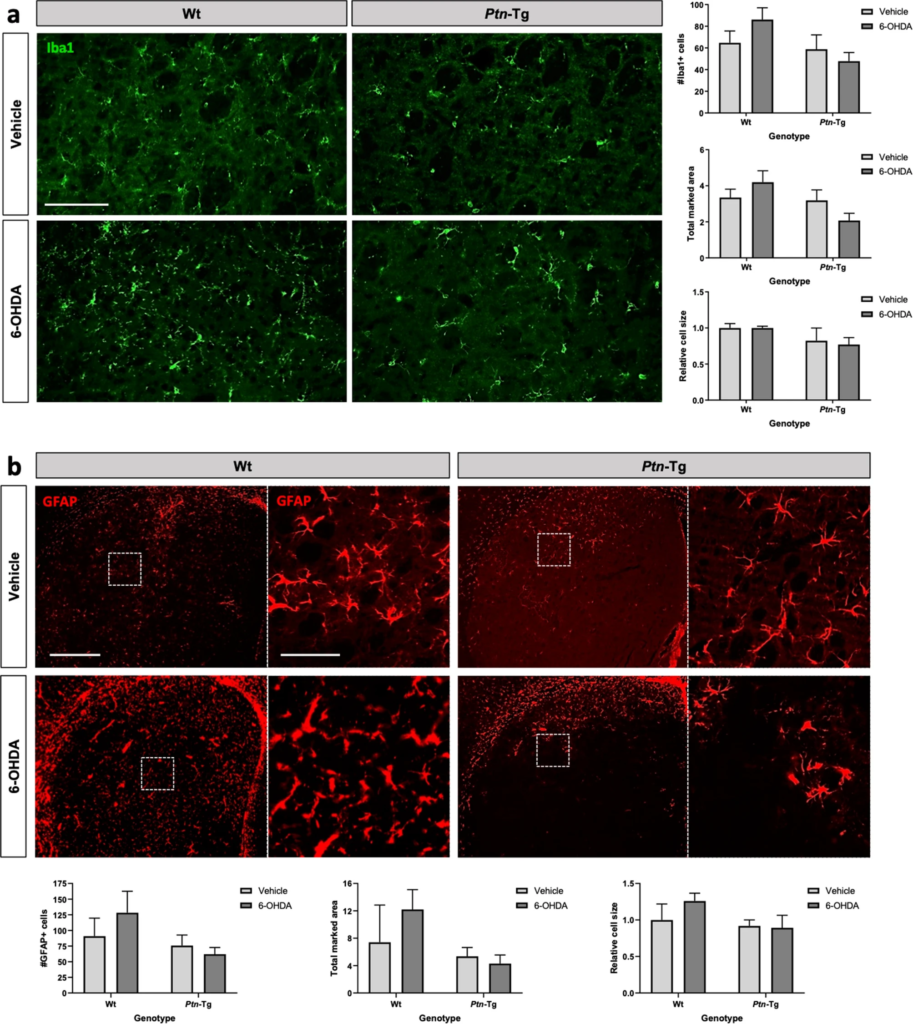
Effects of 6-OHDA on microgliosis and astrocytosis in the striatum of Wt and Ptn-Tg mice. Photomicrographs are representative from Iba1-immunostained (a, green) and GFAP-immunostained (b, red) striatal sections of vehicle and 6-OHDA-injected size of Wt mice and Ptn-Tg mice. Graphs represent quantification of data (mean ± S.E.M) obtained from the counts of Iba1+ cells, total Iba1 marked area and relative cell size (a) and from the counts of GFAP+ cells, total GFAP marked area and relative cell size (b) in the striatum. Scale bar in (a) = 100 μm; Scale bar in (b) = 500 μm (magnification image in dashed line, scale bar = 100 μm).
In summary, the data suggest that PTN overexpression prevents the dopaminergic neurotoxicity induced by a low dose of 6-OHDA that does not cause alterations in motor functions or significant glial responses.
Metabolomics analysis of Ptn-Tg and Wt mice
In general, the analysis performed revealed more significant metabolomic alterations associated with the genotype (Ptn-Tg vs. Wt) within the same treatment than between treatments (6-OHDA vs. vehicle) within the same genotype. The metabolites found significantly altered in the striatum of 6-OHDA-injected Wt mice compared with vehicle-injected Wt mice are listed in supplementary Table 1. The metabolites found significantly altered in the striatum of 6-OHDA-injected Ptn-Tg mice compared with vehicle-injected Ptn-Tg mice are listed in supplementary Table 2. These were mainly polar and small metabolites that did not belong to any particular metabolic group. Further specific analysis for small polar compounds should be carried out in order to confirm the nature of these metabolites and reveal their possible implications in this PD model.
In contrast, we found more profound differences between genotypes, suggesting an important role of cerebral PTN levels in the response to the dopaminergic toxic. The metabolites found significantly altered in the striatum of 6-OHDA-injected Ptn-Tg mice compared with 6-OHDA-injected Wt mice are listed in Table 1. Besides, Fig. 4 shows the OPLS-DA regression model together with the cross-validation scores representation, which shows how these two sets of mice are grouped according to their metabolic profiles. The metabolites found significantly altered in the striatum of vehicle-injected Ptn-Tg mice compared with vehicle-injected Wt mice are listed in Table 2. Similarly, Fig. 5 shows the OPLS-DA regression model together with the cross-validation scores representation, showing the grouping of the mice according to their metabolic profiles in the two models studied. Interestingly, these metabolites coordinately varied in groups (Fig. 6). The main groups of metabolites that varied between 6-OHDA-injected Ptn-Tg and Wt mice belonged to the groups of organic acid with relatively long chains, carnitines, cholesterol derivatives, diacylglycerols (DG), phosphatidylcholines (PC), phosphatidylethanolamines (PE) and phosphatidylserines (PS) (Fig. 6a; Table 1). The altered metabolites between vehicle-injected Ptn-Tg and Wt mice belonged to the groups of carnitines, groups of organic acids with relatively long chains, DG, PC, phosphatidylglycerols (PG), sphingomyelins (SM), phosphatidylinositols (PI) and glucosylceramides (GlcCer) (Fig. 6b; Table 2).
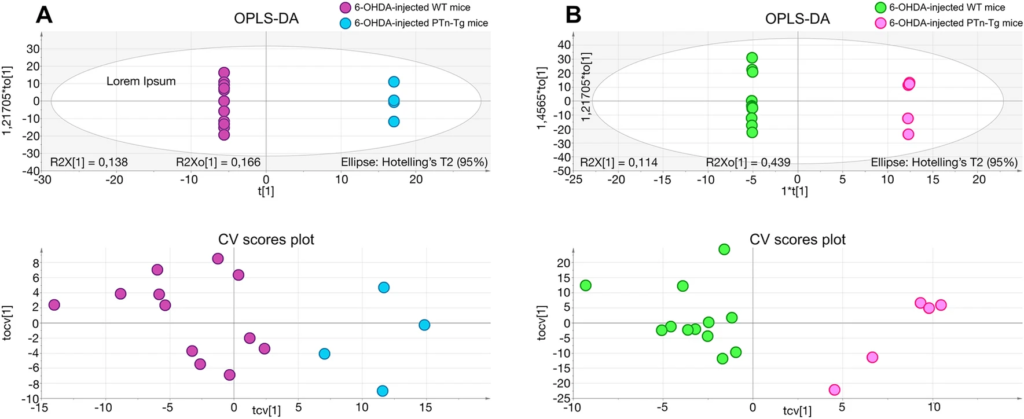
OPLS-DA regression models and cross-validated (CV) score plots showing differentiation of the metabolic profiles between 6-OHDA-injected Wt mice and 6-OHDA-injected Ptn-Tg mice in positive (a, R2 = 0.938, Q2 = 0.699) and negative (b, R2 = 0.855, Q2 = 0.796) ionization modes.
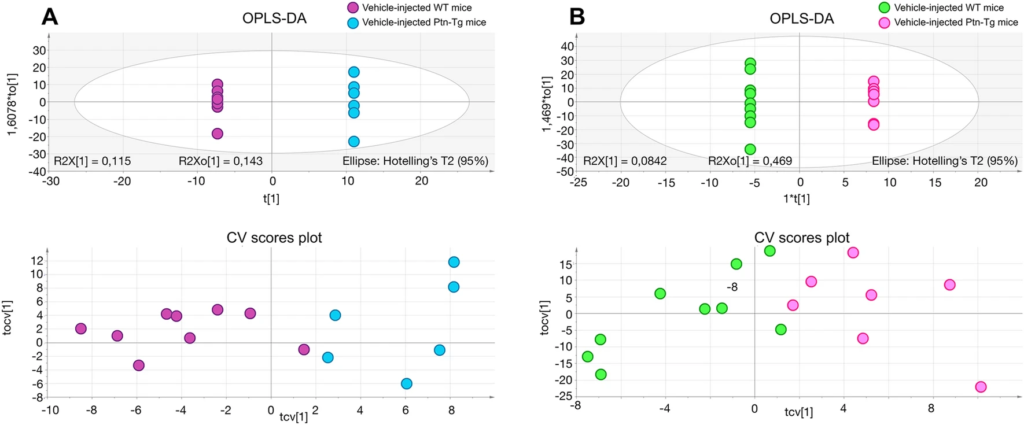
OPLS-DA regression models and cross-validated (CV) score plots showing differentiation of the metabolic profiles between vehicle-injected Wt mice and vehicle-injected Ptn-Tg mice in positive (a, R2 = 0.927, Q2 = 0.686) and negative (b, R2 = 0. 968, Q2 = 0.605) ionization modes.
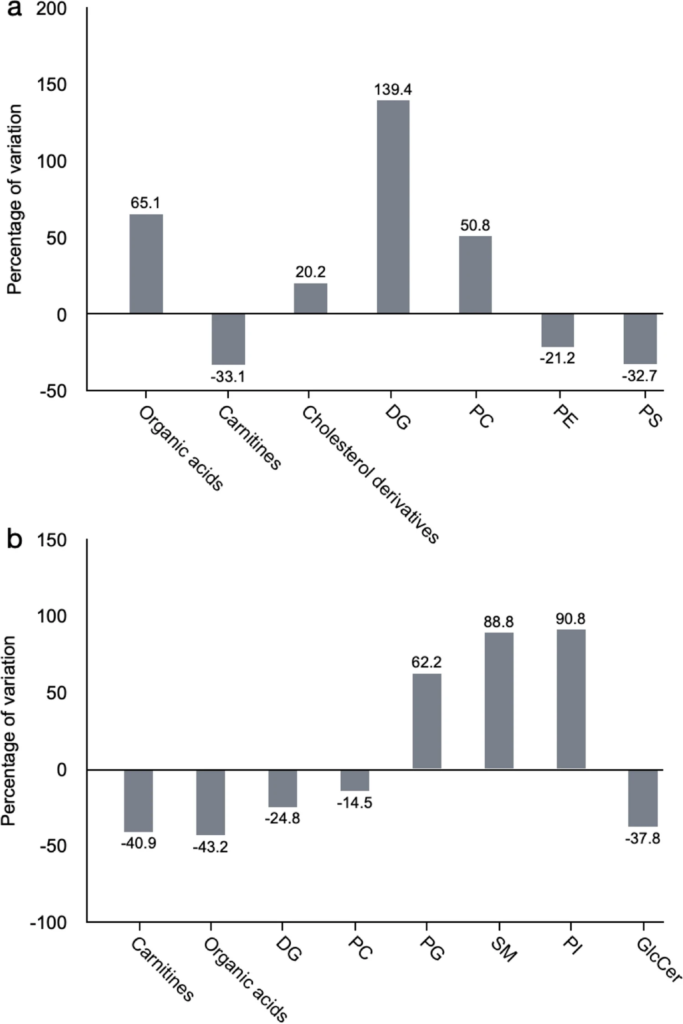
Average percentage of variation for relevant groups of metabolites changed in the different studied groups. (a) Percentage of variation of metabolic groups between Ptn-Tg mice and Wt mice injected with 6-OHDA. (b) percentage of variation of metabolic groups between Ptn-Tg mice and Wt mice injected with the vehicle. DG: Diglyceride, PC: Phosphatidylcholine, PE: Phosphatidylethanolamine, PS: Phosphatidylserine, PG: Phosphatidylglycerol, SM: Sphingomyelin, PI: Phosphatidylinositol.
The metabolomics data has been deposited to the Metabolomics Workbench repository (available at https://www.metabolomicsworkbench.org) with the dataset identifier ST001810.
Discussion
Previous studies supported that PTN exerts neurotrophic effects on dopaminergic neurons, suggesting an important role of this cytokine in PD39. Previously, it was shown that striatal and nigral PTN over-expression provides neuroprotection against 6-OHDA in rats16,17. This effect was extended in the present work by uncovering the correlation between the effects caused by PTN over-expression in the 6-OHDA-injured mouse striatum with relevant changes in striatal phosphatidylcholines, phosphatidylethanolamines, phosphatidylserines, phosphatidylinositols and diglycerides.
Assessing the efficacy of neuroprotective strategies in 6-OHDA models with a full (> 90% striatal dopaminergic denervation) or partial (60–70%) lesion is less than ideal40, probably because of the severity of the injury and the high variability of the resulting symptoms. Here, using a low dose of 6-OHDA (5 µg) to cause a partial lesion in mice, we show that the striatal dopaminergic denervation caused by 6-OHDA is prevented in mice with transgenic PTN overexpression in the brain. In agreement with previous studies with similar levels of striatal denervation41,42, the TH loss caused by this dose of 6-OHDA in Wt mice did not cause significant motor alterations. Moreover, 6-OHDA-induced loss of dopaminergic cells in the substantia nigra seems to be reduced in Ptn-Tg mice compared to Wt mice. It has to be noted that we observed a slightly reduced number of TH+ cells in the substantia nigra of vehicle-injected Ptn-Tg mice compared to Wt mice. Thus, we cannot rule out the possibility that the effect of 6-OHDA is partially masked in Ptn-Tg mice when compared with vehicle-injected Ptn-Tg mice. Although the decrease in TH+ cells in the SNpc is widely considered as dopaminergic neurons loss43,44, in some cases features of apoptosis or necrosis have not been detected45,46, suggesting that lack of TH does not necessarily imply cell death. These differences may be related with the dose and timeline of the study. Therefore, it cannot be assumed that the data presented here represent survival of dopaminergic neurons. However, based on the known neurotrophic actions of PTN on dopaminergic neurons4,17,47,48, it seems reasonable to suggest a neuroprotective role of this cytokine in the SN. Overall, the data demonstrate that PTN limits the nigrostriatal dopaminergic injury in this mouse model of PD.
Striatal gliosis is a hallmark of PD and, in general, of the neuroimmune response triggered by neurotoxins such as 6-OHDA. Besides, PTN has been recently shown to modulate these glial responses after different insults including LPS21,37 and amphetamine6,36 administrations. Thus, we aimed to test if the protective effects of PTN on dopaminergic neurons are paralleled by a regulation of potential glial responses after the administration of a low dose of 6-OHDA. We did not observe relevant responses of microglia and astrocytes to the striatal injection of 6-OHDA in either genotype, suggesting that the striatal TH loss was not accompanied by generalized glial responses. It is important to note that a limitation of this study is that glial responses were only assessed in the striatum to be able to correlate these responses with potential changes in the striatal metabolome. However, testing the neuroinflammatory response in the SN in this experimental model should be of interest for future studies. Another limitation is that we only assessed nigrostriatal injury and glial responses at one time point, ten days after 6-OHDA intrastriatal injection. Thus, it is possible that TH loss or delayed glial changes are more pronounced at a later time point.
The cause of striatal degeneration of dopaminergic terminals is not well known, but increasing evidence suggests a differential astrocytes participation in this process among other factors. Astrocytes can promote or prevent neuronal damage, and the loss of the balance between both opposing actions could be critical for the onset and progression of PD49,50,51,52. The use of suitable animal models could enhance the understanding of the role of striatal denervation and astrocytosis in PD. Thus, in order to improve our understanding of altered metabolic pathways and metabolites in PD, we used the Ptn-Tg mouse as a genetic model of reduced 6-OHDA-induced striatal denervation compared to control, Wt mice. We hypothesize that data from metabolomics studies using this model could result in novel treatments for alterations of metabolic pathways in PD and/or could be used for disease prediction and early diagnosis since our model was performed in the asymptomatic phase.
This study reveals interesting metabolites that significantly discriminate the more injured 6-OHDA-injected Wt striatum and the more protected 6-OHDA-injected Ptn-Tg striatum. Notably, we detected groups of metabolites, mostly corresponding to phospholipids, together with a wide variety of organic acids and carnitines, whose trends were opposite in both groups (Fig. 6a). The fact that entire groups of phospholipids tend to vary together is very significant because α-Synuclein is a lipid-binding protein that interacts with phospholipids and fatty acids53,54.
We found increased levels of 8 forms of phosphatidylcholine (PC) in the striatum of 6-OHDA-injected Ptn-Tg mice compared to Wt mice. Interestingly, this pattern was opposite in vehicle-injected mice. PC is the most abundant phospholipid in cellular membranes55. Decreased levels of PC containing polyunsaturated fatty acyl side chains have been shown in PD brains56,57,58. Importantly, injection of 6-OHDA in rats causes decreases in most PC species in the nigrostriatal circuits59. Our data point to a role of PTN in the increase of PC species after 6-OHDA injection to Ptn-Tg mice compared to Wt mice, which correlated with the significant reduction of dopaminergic injury in their striata. Since PC is known to be involved in neuronal differentiation, neurite outgrowth, and axonal elongation60, it is tempting to hypothesize that the neurotrophic effects of PTN on dopaminergic neurons can be mediated, at least partially, by the increase of PC species.
After PC, the next most abundant group of phospholipids is the group of phosphatidylethanolamines (PE). Five PE species were found decreased in the striatum of 6-OHDA-injected Ptn-Tg mice compared to Wt mice, whereas two species of PE were increased. Decreases in multiple PE species were observed in brains of PD patients56,61. In contrast, increased PE synthesis in the substantia nigra of PD patients has been suggested during the course of PD development62. In general, increased PE synthesis contributes to the rescue of neurons63,64. Our data suggest that some PE (such as PE (39:6) and PE-Cer (32:1)) might be critical in the reported neuroprotective effects of this phospholipid since they are the only PE increased in the striatum of the less vulnerable genotype to 6-OHDA effects, the Ptn-Tg mouse, while the major trend of the majority of these lipids is to decrease in the 6-OHDA-injected Ptn-Tg mice compared to Wt mice.
Eight species of phosphatidylserines (PS) were found decreased in the striatum of 6-OHDA-injected Ptn-Tg mice compared to Wt mice, whereas only one was increased. Decreased PS levels have been observed in 6-OHDA-treated SH-SY5Y cells65, a cellular model of PD, and in plasma from PD patients66. PS is a less abundant membrane phospholipid but is a relevant precursor of mitochondrial PE. Accordingly, as it happened with PE, PS increases and elevated activity of phosphatidylserine synthase, the enzyme responsible for PS synthesis, have been found in the SN of PD patients62. Our results support this hypothesis since general decreases of PS species were found in the less vulnerable genotype to 6-OHDA-induced striatal dopaminergic injury.
Five species of phosphatidylinositols (PI) were found increased in the striatum of vehicle-injected Ptn-Tg mice compared to Wt mice, suggesting that increased PTN brain levels correlate with increased PI levels in basal conditions. In contrast, we did not find significant changes in the striatum of 6-OHDA-injected Ptn-Tg mice compared to Wt mice. The PI total lipid class is significantly reduced in the SN of PD patients58, suggesting an implication of PI depletion in PD. Our data support the possibility that the increases of PI species found in basal conditions in Ptn-Tg mice may play a role in the lower dopaminergic injury found in this genotype after 6-OHDA insult.
Six species of diglycerides (DG) were decreased in the striatum of vehicle-injected Ptn-Tg mice compared to Wt mice, whereas only one was increased. In contrast, we found increases of two DG species in the striatum of 6-OHDA-injected Ptn-Tg mice compared to Wt mice, whereas only one was found decreased. The relevance of these changes needs to be clarified. However, variations in DG species were expected and validate our studies because DGKQ, a diacylglycerol kinase controlling cellular DG, is a designated PD risk factor67,68.
Our data confirm previous findings of known phospholipids that are associated to neurodegeneration, as observed in animal models of PD. Besides, we provide new evidence supporting that the potentiation of PTN signaling may be a novel target for PD, particularly in early stages of the disease since our model was performed in an asymptomatic phase. New lipid-related PD drug candidates emerge from this study, which suggests their significant role in PD. The data presented here support the increasingly recognized “lipid cascade” in PD.